








1. 서론
활성산소종(reactive oxygen species, ROS)은 생리적 수준에서 신호전달, 산화환원 항상성 유지, 세포 대사 조절 등 다양한 생리 기능을 조절하는 필수적 분자로 작용한다(Jomova 등, 2025). 그러나 ROS가 과도하게 축적되면 DNA, 단백질, 지질 등 주요 생체분자를 손상시키는 산화 스트레스를 유발하며, 이로 인한 염증 반응의 악화는 여러 만성 질환의 발병과도 밀접한 관련이 있다(Chatterjee, 2016). 염증 반응은 상처 혹은 외부 이물질 침입 시 신체 방어를 위해 우선적으로 발생하는 선천성 면역 반응으로, 작용과 기간에 따라 급성염증과 만성염증으로 구분된다(Ferrero-Miliani 등, 2006). 급성염증은 외부 자극에 의해 대식세포가 활성화되어 염증매개물질인 cytokine과 nitric oxide (NO), prostaglandin E2 (PGE2), chemokine 등을 생성하는 과정으로 시작된다. 이후 활성화된 백혈구와 대식세포가 이물질을 제거하고 조직을 재생함으로써 염증 반응이 종료된다(Medzhitov, 2008). 이러한 급성염증 반응은 신체 방어에 필수적이지만, 염증매개물질이 과도하게 분비될 경우 혈관 확장에 의한 급성 패혈성 쇼크를 유발하거나, 면역세포의 지속적인 과활성으로 인해 만성염증 혹은 면역과민반응으로 발전할 수 있다(Iwalewa 등, 2007). 이는 류마티스 관절염 등 다양한 만성염증질환 및 자가면역질환의 원인이 될 수 있으므로, 염증 반응의 적절한 조절은 생명현상 유지에 매우 중요하다. 면역세포에는 대식세포와 백혈구, 단핵구, 비만세포 등이 있으며, 그중 모든 조직에 분포한 대식세포는 염증의 유발 원인을 가장 먼저 인식한다. 대식세포는 이를 제거하기 위해 염증매개물질을 생성하고, 다른 면역세포의 활성화 및 주화성을 유도하여 염증 원인을 제거하고 염증 부위의 복구를 돕는다(Fujiwara와 Kobayashi, 2005). 특히, 그람음성균의 세포벽 성분인 lipopolysaccharide (LPS)가 대식세포 표면의 toll-like receptor 4에 결합하면, nuclear factor-kappa B (NF-κB) 신호전달 경로가 활성화된다. 활성화된 NF-κB는 핵으로 이동하여 TNF-α와 IL-6 등 염증성 cytokine 및 inducible NO synthase (iNOS)와 cyclooxygenase-2 (COX-2) 등 염증성 효소의 발현을 촉진함으로써 염증 반응을 유도한다(Baldwin, 1996; Kang 등, 2011). 염증이 과도하게 유발될 경우 NOS의 활성은 급격히 증가하며, 이에 따라 다량의 NO가 생성되어 염증 반응을 더욱 증폭시키는 것으로 알려져 있다(Aleksandrowicz 등, 2025). 이러한 NOS는 neuronal NOS (nNOS)와 endothelial NOS (eNOS), iNOS로 구분된다(Ryszkiewicz 등, 2025). 이 중 eNOS와 nNOS가 생리적 수준에서 혈관 항상성 및 세포 기능 조절에 기여하는 반면(Antosova 등, 2017), iNOS는 LPS나 염증성 cytokine과 같은 자극에 의해 높은 수준으로 발현되어 다량의 NO를 지속적으로 생성하기 때문에 전통적으로 병리적 특성을 나타내는 효소로 간주되어 왔다(Nussler 등, 1992). COX는 COX-1과 COX-2 두 가지 동형 효소로 구성되며, COX-1은 정상적인 생리 기능에 관여하는 구성적 효소인 반면, COX-2는 염증성 cytokine이나 성장인자에 의해 유도되어 염증성 prostaglandin을 생성하는 데 핵심적인 역할을 한다(Droździk 등, 2025).
가래나무(Juglans mandshurica Maxim.)는 호두과(Juglandaceae)에 속하는 낙엽성 교목으로, 중국과 시베리아 및 한반도 중 ․ 북부 지역의 해발 100-1,500 m 산지에 자생하는 종이며, ‘추자목(楸子木)’이라고도 하고, 그 열매는 ‘추자(楸子)’로 불린다(Kwon 등, 2008). 가래나무의 열매는 구형, 난형 또는 타원형에 끝이 뾰족하며, 일반적으로 길이는 3.5-7.5 cm, 지름은 3-5 cm이다(Luan 등, 2021). 또한 가래나무는 약용 식물로, 전통적으로 식도암, 위암, 심장암, 폐암 치료에 사용되어 왔다(Park 등, 2012). 가래나무는 현재까지 다양한 약용 부위(뿌리, 줄기, 나무껍질, 가지, 잎 및 미숙 과일)에 대한 식물화학적 연구를 통해 quinones, phenols, flavonoids, lignans, terpenes, diarylheptanoids, steroids 등을 포함한 다양한 화합물을 분리 및 규명하였다(Guo 등, 2017). 그중 naphthoquinones와 flavonoids, terpenoids, diaryl heptanes가 주요 생리활성 물질로서 광범위하게 연구되었으며(Yao 등, 2016), 이는 전통 중국 의학에서 피부염, 암 등의 치료를 위해 사용된 것으로 확인되었다(Sun 등, 2011). 한편, 가래나무의 열매에는 ellagic acid, juglanin, juglone 등과 같은 다양한 활성 성분이 함유된 것으로 보고되었으며, 이들 화합물은 항종양, 항염증, 항산화 및 항미생물 활성을 나타내는 것으로 알려져 있다. 이러한 성분 조성은 가래나무 열매가 잠재적인 생리활성 소재로서 활용될 가능성을 시사한다(Choi 등, 2024).
현재까지 가래나무의 뿌리(Diao 등, 2017), 잎(Park와 Oh, 2013), 껍질(Qi 등, 2024) 등은 항산화 및 항염증에 대한 효능 연구가 보고되어 있지만, 가래나무의 열매에 관한 생리활성 및 항염증 효능 연구는 상대적으로 제한적인 실정이다. 특히 열매를 대상으로 한 기존 연구들은 주로 메탄올 또는 에탄올 추출물을 중심으로 수행되어 왔다(Luan 등, 2021). 이에 본 연구에서는 가래나무 열매 열수 및 70% 에탄올 추출물을 이용하여 항염증 활성 검증을 통해 기존 연구와는 다른 추출 조건에서의 생리활성 검증을 수행하여 천연 소재로서의 활용 가능성을 확인하고자 하였다.
2. 재료 및 방법
가래나무 열매는 참두리약초(참두리, Seoul, Korea)에서 구입한 후 깨끗하게 세척하였고, 60°C에서 12시간 이상 열풍을 이용하여 건조한 뒤, 대용량 믹서기로 파쇄하여 시료로 이용하였다. 가래나무 열매 열수 추출은 시료에 3차 증류수를 10배량(w/v) 첨가한 후, 환류 냉각을 거쳐 100°C에서 3시간 동안 침지시켰다. 70% 에탄올 추출은 시료에 70% 에탄올을 10배량(w/v) 가한 뒤, 24시간 동안 교반하여 추출하였다. 이후, 부직포를 사용하여 상등액과 침전물을 분리하였으며, 이와 같은 과정을 총 3회 반복하여 추출하였다. 이를 통해 얻은 가래나무 열매 열수 및 70% 에탄올 추출물의 상등액은 정성여과지(Whatman Grade 5, 4, 2)와 진공펌프를 사용하여 침전물과 상등액을 분리하였다. 여과가 완료된 가래나무 열매 열수 및 70% 에탄올 추출물을 rotary vacuum evaporator를 사용하여 용매를 제거하였고, 농축된 추출물은 동결건조하여 powder 형태로 제조하였다. 완성된 시료는 −20°C에 보관하며 본 실험에 사용하였다.
세포 배양 및 실험에 사용한 fetal bovine serum (FBS), penicillin/streptomycin, dulbecco’s modified eagle medium (DMEM), trypsin은 Thermo Scientific HycloneTM (Logan, UT, USA)에서 구입하였으며, 세포 독성 및 NO 생성 억제 활성을 측정하기 위해 사용한 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT), dimethyl sulfoxide (DMSO), LPS, griess reagent 등은 Sigma Chemical Co. (St. Louis, MO, USA)에서 구입하였다. 단백질 정량에 필요한 시약인 protease & phosphatase single-use inhibitor cocktail 100X는 Thermo Scientific HycloneTM (Logan, UT, USA)에서 구입하였으며, RIPA Buffer는 Sigma Chemical Co. (St. Louis, MO, USA)에서 구입하였다. 단백질 발현을 확인하기 위해 사용된 1차 항체인 β-actin, COX-2와 2차 항체인 anti-mouse는 Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA)에서 구입하였고, 1차 항체인 iNOS와 2차 항체인 anti-rabbit은 Cell Signaling Technology Inc. (Danvers, MA, USA)사 제품을 사용하였다. mRNA 발현 측정을 위한 실험에 사용된 TRIzol® reagent는 Thermo Scientific Ambion® (Austin, TX, USA)에서 구입하였고, Ethidium Bromide Solution (EtBr)은 Bio-Rad (Hercules, CA, USA)에서 구입하여 사용하였다. 또한, epigenTM cDNA Syn. dT15 Master Mix, 4x와 epigenTM Hot Start Premix, 2x는 EPIGEN Co., Ltd. (Cheongju, Korea)에서 구입하였으며, primer인 GAPDH, iNOS와 COX-2는 Bionics (Seoul, Korea)에서 구입하여 실험에 사용하였다. 염증성 cytokine을 측정하기 위해 사용한 TNF-α와 IL-6 enzyme-linked immunosorbent assay (ELISA) kit는 R&D Systems, Inc. (Minneapolis, MN, USA)에서 구입하였다.
실험에 사용된 기기는 vortex mixer (SI-0256, Scientific Industries, Inc., Bohemia, NY, USA), rotary evaporator (EYELA N-1000, EYELA, Tokyo, Japan), freeze dryer (FD5508, ILShin Lab Co., Ltd., Seoul, Korea), centrifuge (MF-300, Hanil Scientific Inc., Gimpo, Korea), micro refrigerated centrifuge (Smart-R17, Hanil Scientific Inc.), microplate reader (SPECTROstar Nano, BMG LABTECH, Ortenberg, Germany), nano drop (Q5000, Quawell Technology, Inc., San Jose, CA, USA), CO2 incubator (MCO-18AC, Panasonic Healthcare Co., Kadomashi, Osaka, Japan), ChemiDocTM Imaging System (BIO-RAD, Hercules, CA, USA)을 사용하였다.
본 실험에 사용된 세포주는 미국세포주은행(American Type Culture Collection; ATCC, Manassas, WV, USA)에서 구입하였으며, RAW 264.7 (TIB-71) 대식세포를 이용하였다. 세포는 DMEM에 FBS (10%)와 penicillin/streptomycin (100 U/mL, 1%)을 혼합한 배지에서 세포가 성장하는 속도에 맞춰 계대 배양하였다. 모든 배양 과정은 37°C, 5% CO2 조건의 incubator에서 이루어졌다.
세포 생존율은 Carmichael 등(1987)의 방법을 참고하여 측정하였다. 먼저, 96 well cell culture plate에 5×103 cells/well 양의 세포를 100 μL씩 분주하여 incubator (37°C, 5% CO2 조건)에서 24시간 동안 배양하였다. 이후 세포에 농도별(5, 10, 50, 100, 500, 1,000 μg/mL)로 시료를 처리하고 동일한 조건에서 다시 24시간 동안 배양을 진행하였다. 배양이 끝난 뒤, 세포에 2.5 mg/mL 농도로 제조한 MTT 시약을 각 well에 40 μL씩 첨가하여 4시간 동안 반응시켰다. 반응 후 상층액을 제거하고, 각 well에 DMSO 100 μL씩 가하여 실온에서 10분간 shaking한 다음 microplate reader를 이용해 540 nm에서 흡광도를 측정하였다. 세포 생존율은 시료의 첨가 그룹과 무첨가 그룹의 흡광도에 대한 백분율로 나타내었다.
LPS 자극제로 염증 반응을 유도하여 RAW 264.7 세포가 배양액에 생성시킨 NO양 측정은 Green 등(1982)의 방법을 참고하여 측정하였다. 세포는 6 well cell culture plate에 8×105 cells/well 양의 세포를 1 mL씩 분주하여 incubator (37°C, 5% CO2 조건)에서 24시간 동안 배양하였다. 이후 LPS 무처리 그룹을 제외한 control (LPS 단독 처리 그룹)과 각 농도 구간에 LPS를 처리하여 염증 반응을 유도한 후, 2시간 뒤 추출물을 2.5, 5, 10 μg/mL의 농도로 각각 첨가하여 16시간 동안 추가로 배양하였다. 배양이 끝난 후 상층액을 회수하여 96 well cell culture plate에 100 μL씩 옮기고, griess reagent 100 μL를 더한 뒤 실온에서 10분간 shaking하였다. 반응이 끝난 시료는 microplate reader를 사용해 540 nm에서 흡광도를 측정하였다. NO 생성 억제율은 시료의 첨가 그룹과 무첨가 그룹을 비교하여 본 흡광도에서 백분율 산출로 나타내었다.
iNOS와 COX-2 단백질 발현을 측정하기 위해 100 mm cell culture dish에 1×106 cells/well 양의 RAW 264.7 세포를 1 mL씩 분주하여 incubator (37°C, 5% CO2 조건)에서 24시간 동안 배양하였다. 세포가 안정화된 후, LPS 무처리 그룹을 제외한 control (LPS 단독 처리 그룹)과 각 농도 구간에 LPS를 먼저 처리하였고, 2시간 후 농도별(2.5, 5, 10 μg/mL)로 추출물을 추가하여 16시간 더 배양하였다. 배양이 끝난 뒤, 배지를 제거하고 PBS로 세포를 2번 세척하였다. 이후 RIPA buffer 10 mL에 protease & phosphatase single-use inhibitor cocktail (100X) 100 μL를 첨가해 제조한 lysis buffer로 세포막을 파괴하였다. 용해된 세포는 4°C, 16,110 ×g 조건으로 20분간 원심 분리하여 상층액을 확보하였다. 세포 상층액 내의 단백질 정량은 BCA protein assay kit를 이용해 수행하였으며, 4x laemmli sample buffer와 혼합하여 정량된 단백질 용액 20 μL를 10% sodium dodecyl sulfate acrylamide gel에 loading하여 120 V에서 전기영동하였다. 전기영동으로 분리된 단백질은 transfer 장치를 이용하여 polyvinylidene fluoride membrane으로 이동시켰고, 이후 실온에서 5% skim milk가 용해된 tris-buffered saline & tween 20 (TBST)로 1시간 동안 blocking하였다. 그 후 β-actin, iNOS, COX-2 1차 항체는 1:1,000으로 희석해 4°C에서 overnight 반응시킨 뒤, TBST로 10분 간격으로 3번 세척하였다. β-actin과 COX-2 anti-mouse 2차 항체는 1:2,500으로 희석하여 사용하고, iNOS anti-rabbit 2차 항체는 1:3,000으로 희석하여 사용하였으며, 실온에서 1시간 30분 동안 배양하였다. 이후 TBST로 10분 간격으로 3번 세척하였다. 마지막으로 암실에서 enhanced chemiluminescence 용액으로 반응시킨 후, ChemiDocTM Imaging System을 사용하여 단백질 밴드를 확인하였다.
6 well cell culture plate에 8×105 cells/well 양의 RAW 264.7 세포를 1 mL씩 분주하여 incubator (37°C, 5% CO2 조건)에서 24시간 동안 배양하였다. 배양이 끝난 뒤, LPS를 각 well에 10 μg/mL가 되도록 처리하고, 2시간이 지난 뒤 추출물을 농도별로 첨가하여 16시간 추가 배양을 진행하였다. 배양이 끝난 후 배지를 제거한 뒤 PBS로 두 차례 세척하였으며, RNA 추출을 위해 각 well에 trizol reagent 1 mL를 분주하여 세포를 lysis 하였다. 이후 chloroform 200 μL를 첨가하여 10-15회 부드럽게 섞은 다음, 층이 분리된 것을 확인한 후 4°C, 16,110 ×g으로 20분간 원심분리를 실시하였다. 상층액은 isopropanol 500 μL와 섞어 동일한 조건으로 다시 원심분리하였으며, 이 과정을 통해 RNA pellet을 확보하였다. Pellet을 제외한 상층액을 제거한 뒤, diethyl pyrocarbonate (DEPC) treated water로 희석한 75% 에탄올을 사용해 세척하고 남은 용액을 제거하였다. 이후 pellet에 DEPC treated water를 50 μL씩 가해 용해시킨 다음, total RNA 함량은 nano drop을 사용하여 측정하였다. cDNA 합성을 위해 추출한 RNA와 nuclease free water (NW)를 혼합하여 70°C에서 5분간 반응을 진행한 뒤, 4°C 조건에서 추가로 5분간 반응시켰다. 이후 Syn. dT15 Master Mix를 첨가하여 50°C에서 10분, 95°C에서 5분, 마지막으로 16°C에서 5분간 반응시켜 cDNA를 합성하였다.
항염증 활성을 평가하기 위해 RAW 264.7 세포에서 염증 매개 인자의 mRNA 발현을 RT-PCR로 측정하였다. 합성된 cDNA는 Hot Start Premix와 각 primer를 혼합하여 반응을 준비하였으며, 실험에 사용된 primer sequences는 Table 1에 제시하였다. PCR 조건은 인자별로 다음과 같이 설정하였다. GAPDH는 95°C에서 3분, 95°C에서 10초, 64°C에서 20초, 72°C에서 20초, 72°C에서 5분으로 30 cycles, iNOS는 95°C에서 3분, 95°C에서 10초, 62°C에서 20초, 72°C에서 20초, 72°C에서 5분으로 30 cycles, COX-2는 95°C에서 3분, 95°C에서 10초, 51°C에서 20초, 72°C에서 20초, 72°C에서 5분으로 35 cycles를 실행하였다. 증폭된 PCR 산물은 5% EtBr이 첨가된 1.5% agarose gel에 100 V 조건에서 30분 이내로 전기영동하였으며, 이후 자외선을 조사하여 밴드를 확인하였다.
염증성 cytokine의 생성량을 측정하기 위해 6 well cell culture plate에 8×105 cells/well 양의 RAW 264.7 세포를 1 mL씩 분주하여 incubator (37°C, 5% CO2 조건)에서 24시간 동안 배양하였다. 이후 세포에 LPS를 10 μg/mL의 농도로 처리하고, 2시간이 지난 뒤 농도별(2.5, 5, 10 μg/mL)로 추출물을 첨가하여 16시간 추가 배양을 진행하였다. 배양이 끝난 뒤, 각 well에서 상층액 500 μL를 회수하여 16,110 ×g에서 5분간 원심분리하였고, 다시 상층액 300 μL를 취하여 측정에 사용하였다. 염증성 cytokine인 TNF-α와 IL-6는 각각의 ELISA kit를 이용해 제조사 매뉴얼에 따라 측정하였으며, 생성량은 microplate reader로 450 nm에서 흡광도를 측정하여 분석하였다.
3. 결과 및 고찰
MTT는 살아있는 세포의 미토콘드리아 내막에 존재하는 oxido-reductase의 효소 반응에 의해 환원되어 보라색의 formazan을 형성한다. 생성된 formazan을 유기용매 등으로 용해한 후 발색 강도를 흡광도로 측정함으로써, 세포 생존율을 측정하는 데 활용되는 방법이다(Denizot과 Lang, 1986).
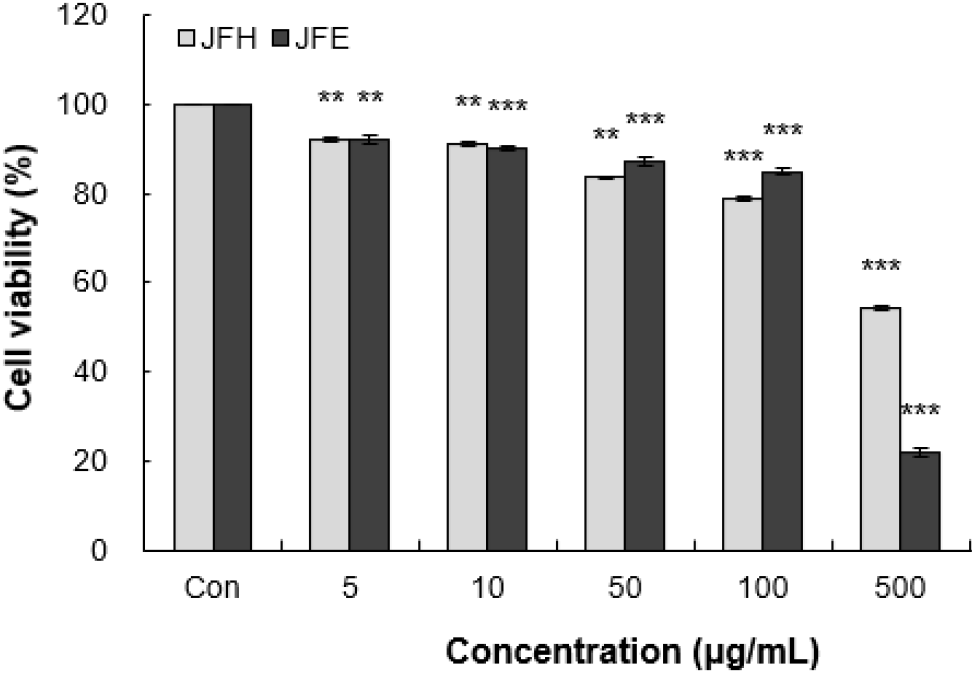
대식세포인 RAW 264.7 세포를 이용해 MTT assay를 수행한 결과, 가래나무 열매 열수 및 70% 에탄올 추출물은 10 μg/mL 이하의 농도에서 각각 91.17%, 90.33%의 세포 생존율을 보였다(Fig. 1). 이러한 결과를 바탕으로 이후 모든 실험은 2.5, 5, 10 μg/mL 농도구간에서 진행하였다.
NO는 지질 친화성이 높은 무색의 기체 자유라디칼로서 세포막을 쉽게 통과하며(Khan 등, 2023), L-arginine이 L-citrulline으로 산화되는 과정에서 생성된다(Knowles와 Moncada, 1994). 염증 과정에서 NO가 과도하게 생성될 경우, 조직 손상을 유발하고 패혈성 쇼크, 만성염증 및 자가면역질환의 원인이 될 수 있다(Miyataka 등, 2002). NO의 생성량은 일반적으로 griess reagent를 이용해 측정하는데, 이는 세포에서 생성된 NO의 산화산물인 아질산염과 반응하여 분홍색의 아조염을 형성하며, 아질산염 농도가 높을수록 더욱 강한 붉은색을 나타낸다(Yucel 등, 2012). 즉, 염증 반응 과정에서 유도된 NO의 양을 색 변화로 정량할 수 있다는 원리를 이용하여, 시료의 NO 생성 억제 활성을 평가하였다. 이에 따라, RAW 264.7 세포에 LPS를 처리하여 염증 반응을 유도한 뒤, 가래나무 열매 열수 및 70% 에탄올 추출물을 2.5, 5, 10 μg/mL 농도로 처리하여 NO의 생성량을 측정한 결과를 Fig. 2에 나타내었다. 그 결과, 가래나무 열매 열수 및 70% 에탄올 추출물은 LPS 단독 처리 그룹 대비 10 μg/mL 농도에서 각각 37.61%, 46.61%의 억제 효과를 나타냈다. 또한, 두 추출물의 억제율은 LPS 단독 처리 그룹 대비 29.44%의 억제율을 보인 무처리 그룹보다 더 높은 수준을 보여, 가래나무 열매 열수 및 70% 에탄올 추출물의 NO 생성 억제 활성이 우수함을 확인하였다.
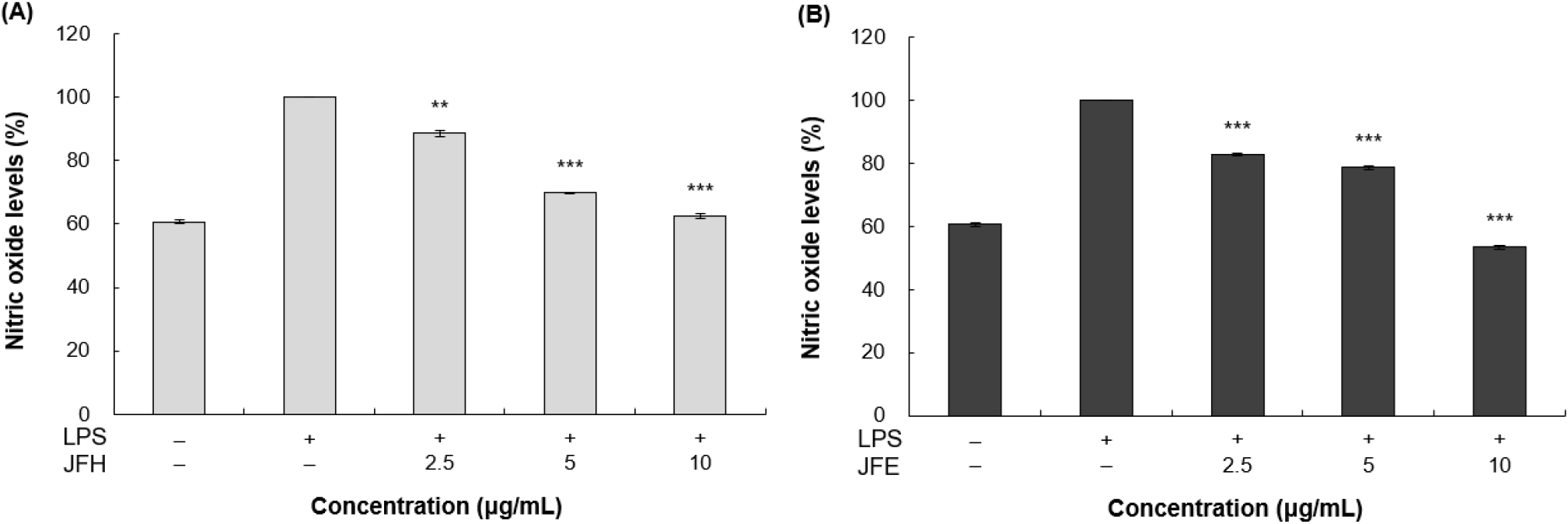
Lee 등(2022)은 아스파라거스 뿌리 열수 추출물이 500 μg/mL 농도에서 26.5%의 NO 생성 억제율을 나타낸다고 보고하였으며, Lee 등(2017)의 채진목 70% 에탄올 추출물이 1,000 μg/mL 농도에서 31.4%의 억제 효과를 보인다고 제시하였다. 이러한 기존 연구와 비교했을 때, 본 연구에서 사용한 가래나무 열매 열수 및 70% 에탄올 추출물은 상대적으로 낮은 농도에서도 더 높은 NO 저해 활성을 보여 우수한 항염증 효과를 지니는 것으로 판단되었다.
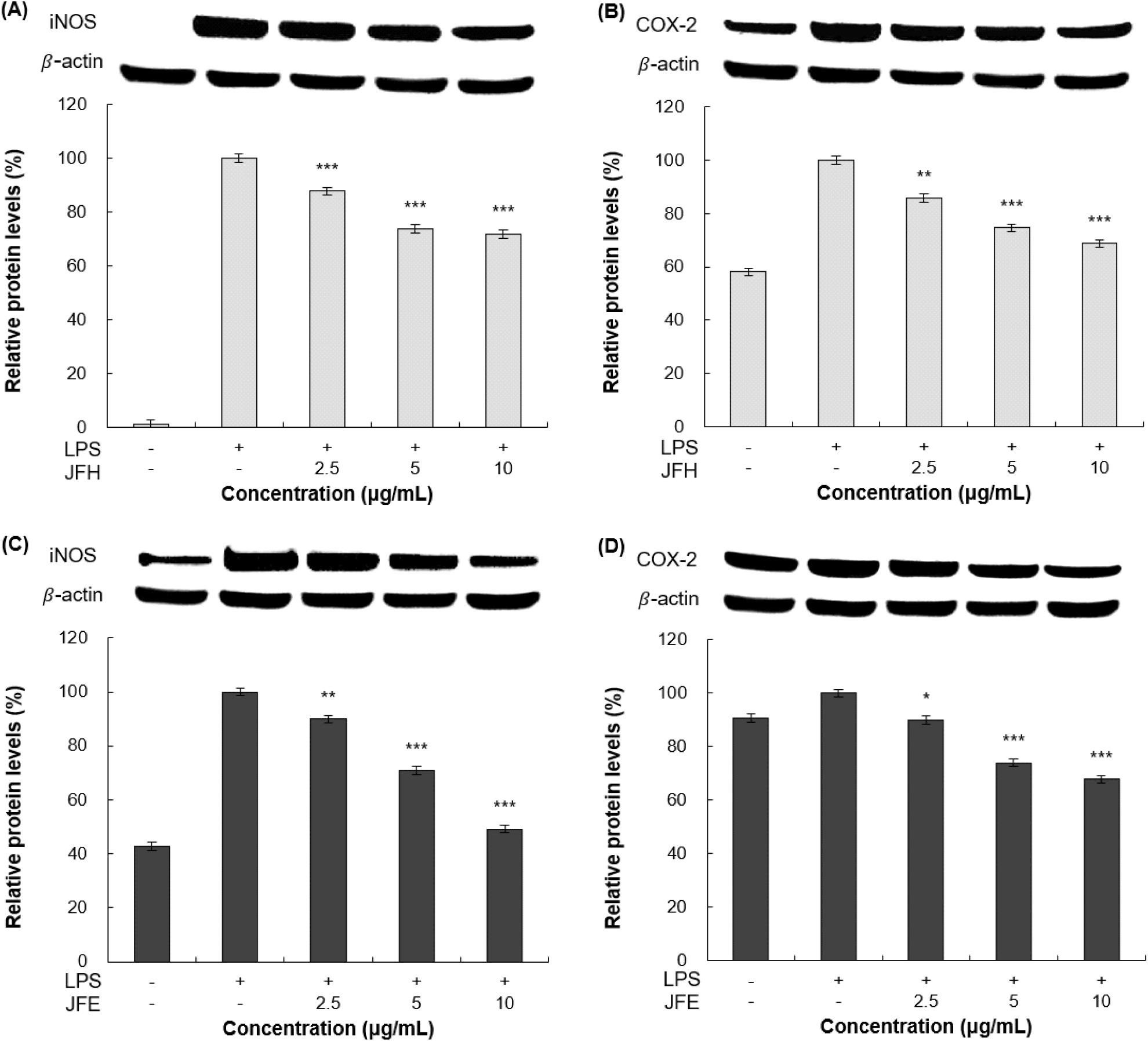
iNOS는 다양한 조직에서 유도적으로 발현되는 효소로, 염증, 저산소증, 뇌 허혈과 같은 병리적 조건에서 그 활성이 과도하게 증가한다(Justo와 Suemoto, 2022). 이 과정에서 생성된 NO는 자유라디칼과 반응하여 과산화질산염 등 강력한 산화 ․ 질소화 스트레스 유발 분자를 형성하며, 이는 세포 독성, 단백질 변성, 지질 과산화 및 조직 손상을 초래해 염증성 질환의 진행에 중요한 역할을 하는 것으로 알려져 있다(Habib 등, 2022). COX-2는 arachidonic acid를 PGE2로 전환하는 반응을 촉매하며(Budiastuti 등, 2025), 특히 급성 및 만성염증 반응에서 통증 유발, 혈관 투과성 증가, 혈관 확장의 염증 반응 주요 매개체로 작용한다(Cerella 등, 2010). 이러한 iNOS와 COX-2의 과도한 생성은 다양한 염증 반응과 조직 손상을 유발하므로, 염증성 질환의 예방과 치료를 위해 이들의 생성을 억제하는 것이 중요하다(Hwang 등, 2016). 따라서 항염 활성을 평가하기 위해 western blot을 통해 염증 인자인 iNOS, COX-2의 단백질 발현 억제 효과를 측정하였다. RAW 264.7 세포에 가래나무 열매 열수 및 70% 에탄올 추출물을 처리한 뒤, 단백질의 발현량을 분석한 결과는 Fig. 3에 나타내었다. β-actin을 housekeeping 인자로 사용하였으며, iNOS와 COX-2의 단백질 발현량이 가래나무 열매 열수 추출물은 10 μg/mL 농도에서 LPS 단독 처리 그룹 대비 각각 28.12%, 31.26%의 저해 효과를 나타내었고, 가래나무 열매 70% 에탄올 추출물은 각각 50.78%, 32.18%의 저해 효과를 보여주었다.
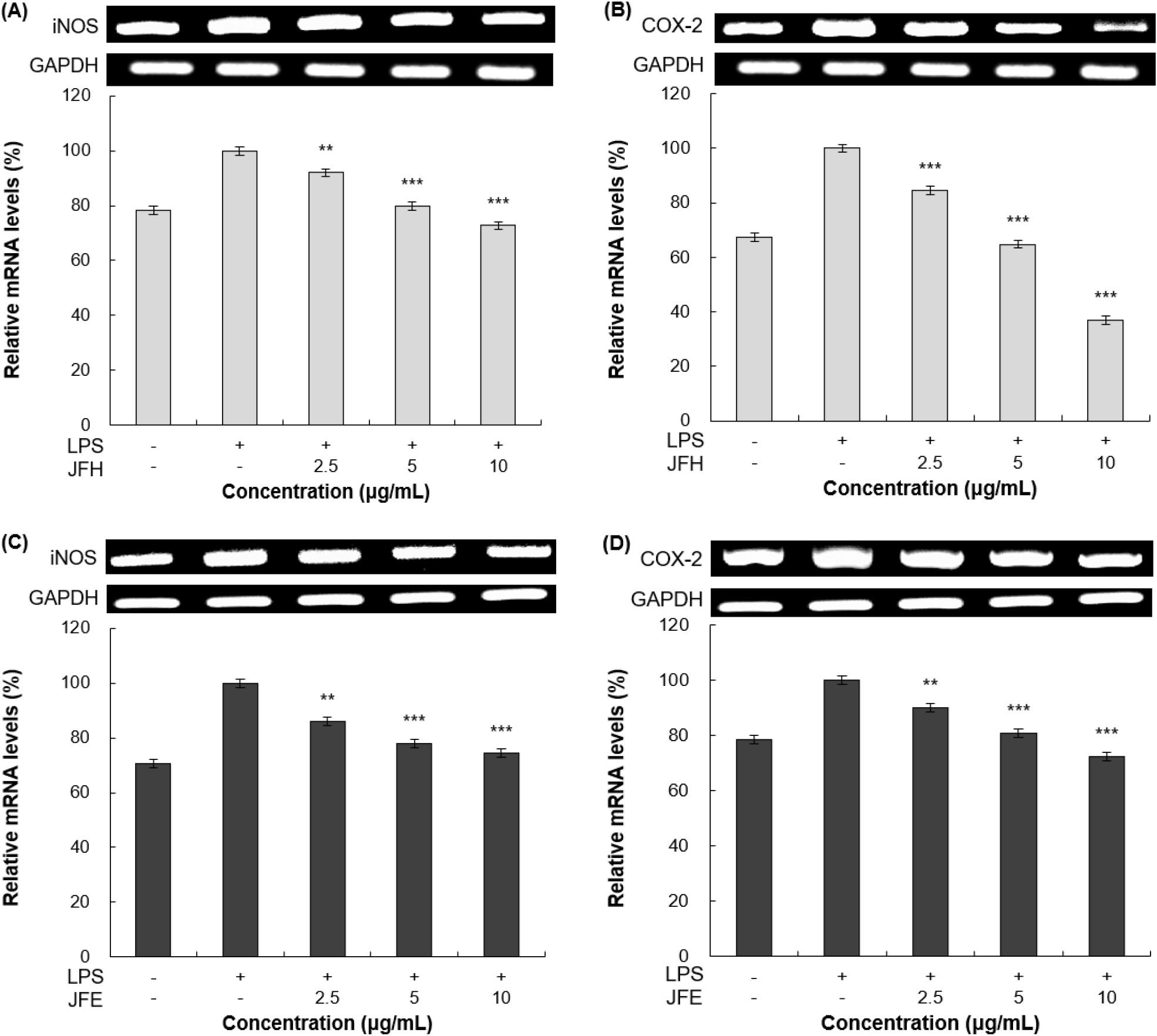
mRNA 수준에서의 iNOS 및 COX-2 발현 억제 효과를 측정하기 위해 RT-PCR을 수행하였다. 가래나무 열매 열수 및 70% 에탄올 추출물을 RAW 264.7 세포에 처리한 뒤 측정한 mRNA의 발현 결과는 Fig. 4에 제시하였다. 먼저, LPS 단독 처리 그룹은 무처리 그룹 대비 뚜렷한 mRNA 발현 증가 양상을 확인할 수 있었다. 이러한 LPS 단독 처리 그룹과 비교하였을 때, 10 μg/mL 농도에서 가래나무 열매 열수 추출물은 iNOS와 COX-2 발현을 각각 27.22%, 62.98% 나타내었고, 70% 에탄올 추출물은 각각 25.55%, 27.47%의 억제 효과를 나타내었다.
TNF-α는 활성화된 대식세포에서 주로 생성되며(Bradley, 2008), 자가분비 및 측분비 방식으로 대식세포의 TNF-α 수용체에 결합하여 NF-κB를 활성화시키고, 이를 통해 TNF-α, IL-6, iNOS, COX-2와 같은 염증성 매개 인자의 발현을 유도한다(Tracey 등, 2008). IL-6는 Janus kinase-signal transducer activator of transcription 신호전달 경로를 통해 T세포와 B세포를 활성화시켜 급성염증을 만성염증으로 전환시키는 데 관여한다(Gabay, 2006). 그러나 TNF-α와 IL-6가 과도하게 분비될 경우 면역계 항상성이 무너져 다양한 세포와 조직의 기능 저하를 초래할 수 있으며, 이는 염증성 장질환, 제2형 당뇨, 류마티스와 같은 만성염증 및 자가면역질환으로 이어질 수 있다. 따라서 이러한 염증성 cytokine인 TNF-α와 IL-6의 생성을 조절할 수 있는 물질은 만성염증 및 자가면역질환의 치료제로 활용될 가능성이 있다(Won 등, 2008).
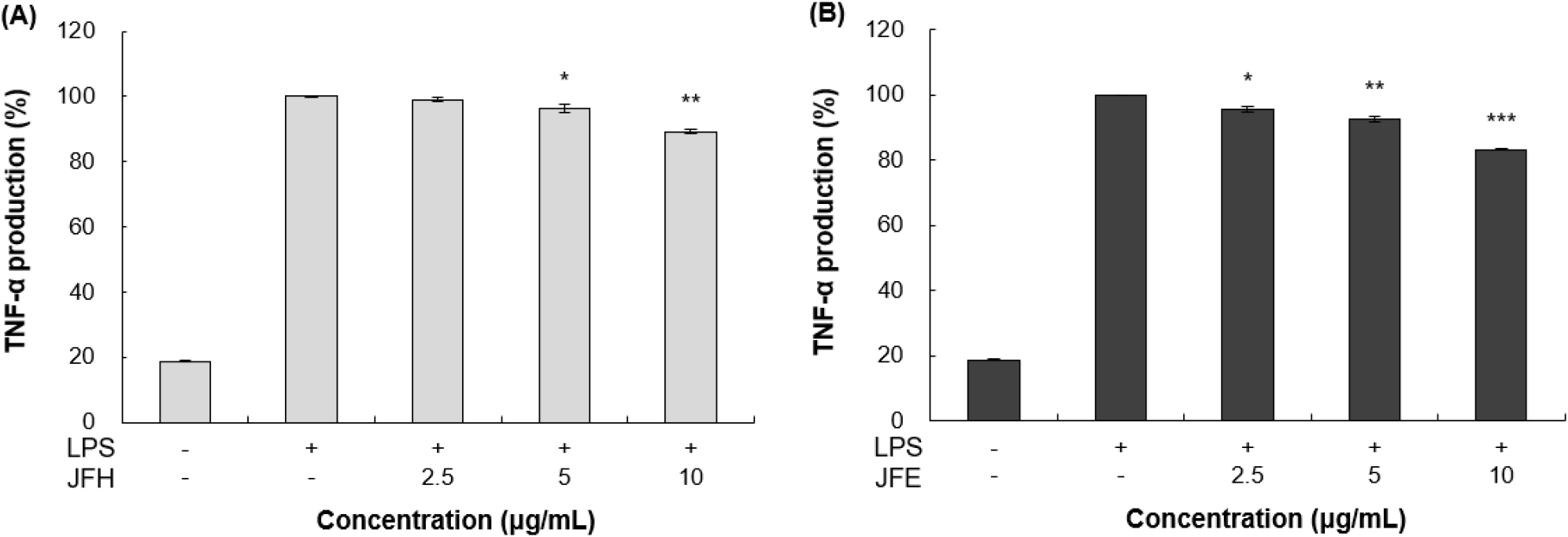
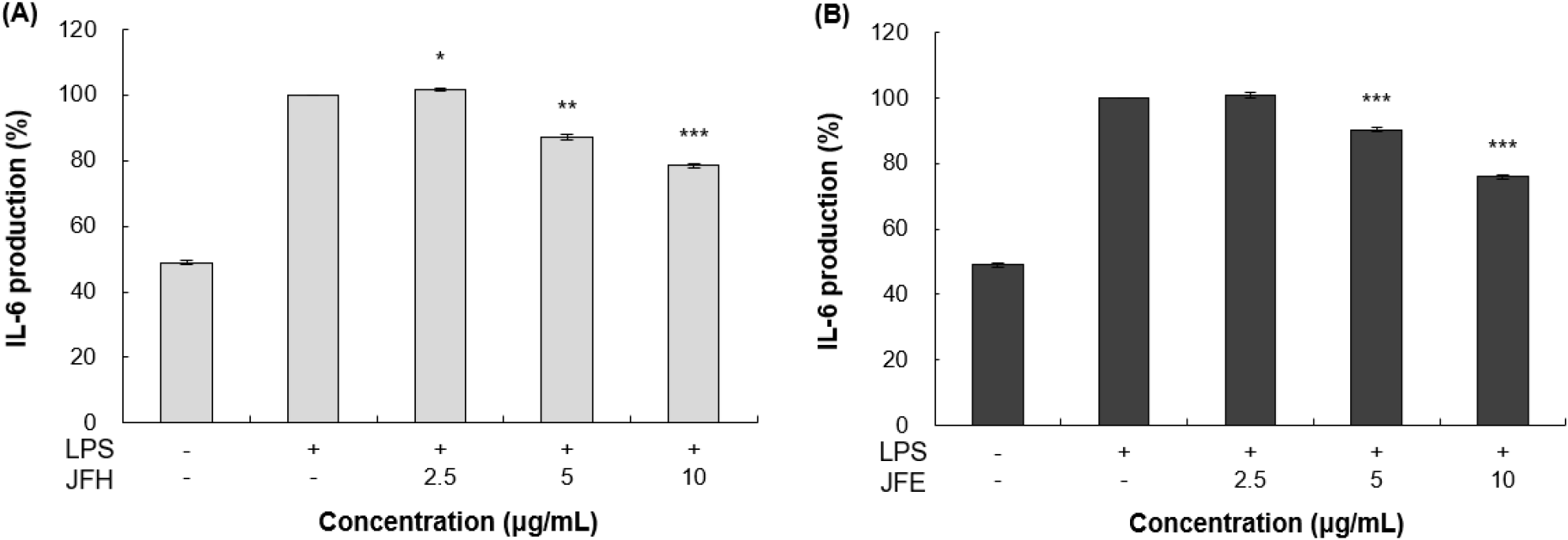
TNF-α 및 IL-6와 같은 염증성 cytokine의 생성 억제를 확인하기 위해 ELISA kit를 이용한 분석을 실시하였다. 가래나무 열매 열수 및 70% 에탄올 추출물을 RAW 264.7 세포에 처리한 뒤, 각 ELISA kit를 사용하여 TNF-α와 IL-6의 발현량을 측정한 결과는 각각 Fig. 5와 Fig. 6에 제시하였다. LPS 단독 처리 그룹은 무처리 그룹 대비 뚜렷한 cytokine 발현 증가 양상을 확인할 수 있었다. 이러한 LPS 단독 처리 그룹과 비교하였을 때, 10 μg/mL 농도에서 가래나무 열매 열수 추출물은 TNF-α와 IL-6를 각각 10.70%, 16.91% 감소시켰다. 또한 가래나무 열매 70% 에탄올 추출물은 동일한 농도에서 각각 21.61%, 24.13%의 억제 효과를 보였다.
본 연구에서는 가래나무 열매의 열수 및 70% 에탄올 추출물을 제조하여 항염증에 대한 효과를 확인하고자 본 실험을 진행하였다. LPS로 활성화된 RAW 264.7 세포에서의 항염증 효능 측정 결과, iNOS와 COX-2의 단백질 및 mRNA 발현을 모두 유의한 수준으로 저해하는 것을 확인하였다. 또한, RAW 264.7 세포에서 염증 반응에 관여하는 TNF-α, IL-6의 생성량을 각각 ELISA kit를 이용하여 측정한 결과, 두 추출물 모두 농도의존적으로 감소하는 것을 보여주었다. 본 연구에서 관찰된 가래나무 열매 열수 및 70% 에탄올 추출물 간 활성 차이는 용매 특성에 따른 추출 성분 조성의 차이와 관련될 가능성이 있으며, 에탄올 추출이 상대적으로 다양한 생리활성 물질의 용출을 유도한다는 기존 보고와 일치하는 경향을 보였다(Park과 Hong, 2014). 결론적으로 가래나무 열매의 열수 및 70% 에탄올 추출물은 염증 발생에 중요한 역할을 하는 TNF-α, IL-6와 같은 염증성 cytokine과 iNOS, COX-2의 발현을 억제하므로 항염 효과를 확인하여 이에 기능성 천연 소재로서 활용 가능성이 있다고 판단된다.
4. 요약
본 실험은 가래나무 열매를 열수 및 70% 에탄올로 추출하여 항염 효과를 알아보고자 하였다. 먼저 RAW 264.7 세포에서의 세포 생존율을 측정한 결과, 가래나무 열매 열수 및 70% 에탄올 추출물은 모두 10 μg/mL 이하의 농도에서 10% 미만의 독성이 측정되었다. 따라서, 이후 실험은 90% 이상의 세포 생존율을 보인 2.5, 5, 10 μg/mL의 농도구간을 설정하여 진행하였다. 항염증 활성은 LPS로 염증 반응을 유도한 RAW 264.7 세포에서 NO 생성 억제 활성을 확인하였으며, LPS 단독 처리 그룹 대비 가래나무 열매 열수 및 70% 에탄올 추출물은 각각 최고 농도인 10 μg/mL에서 37.61%, 46.61%의 억제 효과가 나타났다. 또한, 두 추출물의 억제율은 LPS 단독 처리 그룹 대비 무처리 그룹(29.44%)보다 더 높은 수준을 보여, 가래나무 열매 추출물이 NO 생성을 효과적으로 감소시키는 항염증 활성을 가지고 있는 것을 확인하였다. 염증에 관여하는 대표적 인자인 iNOS 및 COX-2의 단백질 수준에서의 발현량을 측정한 결과, 가래나무 열매 열수 추출물은 LPS 단독 처리 그룹 대비 10 μg/mL 농도에서 각각 28.12%, 31.26%가 억제되었으며, 가래나무 열매 70% 에탄올 추출물은 각각 50.78%, 32.18% 억제됨을 확인하였다. 또한, iNOS와 COX-2의 mRNA 수준에서의 발현량을 측정한 결과, 가래나무 열매 열수 추출물은 LPS 단독 처리 그룹 대비 10 μg/mL 농도에서 각각 27.22%, 62.98%의 억제 효과를 나타내었고, 가래나무 열매 70% 에탄올 추출물은 각각 25.55%, 27.47%의 억제 효과를 나타내었다. TNF-α 및 IL-6와 같은 염증성 cytokine의 생성량은 ELISA kit를 사용하여 측정한 결과, 가래나무 열매 열수 추출물은 LPS 단독 처리 그룹 대비 10 μg/mL 농도에서 각각 10.70%, 16.91% 억제되었으며, 가래나무 열매 70% 에탄올 추출물은 각각 21.61%, 24.13% 억제됨을 확인하였다. 따라서, 가래나무 열매 열수 및 70% 에탄올 추출물은 항염 활성을 나타내어, 기능성 천연 소재로서의 적용 가능성이 높을 것으로 판단된다.