








1. 서론
느타리버섯(Pleurotus ostreatus)은 담자균문 주름버섯목 느타리속에 속하는 버섯으로, 재배가 잘 되는 식용버섯 중 하나이다(Zhang 등, 2002). 우리나라를 비롯한 아시아 지역에서 대량으로 생산되며(Oh 등, 2024), 다양한 요리에 활용되어 왔다. 또한 식용버섯으로서 단백질 ․ 섬유소와 함께 미네랄, 비타민, 키틴, 폴리페놀 등의 성분을 함유하고 있어 영양가가 높으며 다양한 생리활성에 관한 연구도 보고되고 있다(Corrêa 등, 2016; Effiong 등, 2024; Matkovits 등, 2024).
유산균 발효는 식품의 가공 방법 중의 하나로 발효물의 항산화, 항균 및 항염증 효능 강화, 소화 개선, 기능성 물질 생성 등과 관련된 여러 유익한 효능에 대한 연구가 현재까지도 활발하게 진행되고 있다(Chen 등, 2025; Prete 등, 2024; Zhang 등, 2023; Zhao 등, 2023). 유산균은 인간이 이용할 수 있는 유익한 미생물의 한 종류로 유제품을 중심으로 식품 업계에서 광범위하게 활용되고 있으며(Marco 등, 2017), 특히 본 논문에 사용한 Pediococcus pentosaceus는 젖산균의 일종으로 항산화 및 항염 효과와 콜레스테롤 개선 등의 다양한 생리활성을 갖는 것으로 알려져 있다(Qi 등, 2021).
현대사회는 지속적인 경제성장과 의료 발전으로 인해 평균 수명은 증가하였지만 환경 오염 및 불규칙적인 생활 패턴 등의 문제로 질병이 증가하는 추세이며 건강 기능성 식품 등을 통해 이를 예방하고 관리하기 위한 수요 또한 증가하고 있다(Gutiérrez-Grijalva 등, 2024; Kwon 등, 2018; Park, 2018; Song과 Yoon, 2019). 그중 만성 염증과 알레르기 질환은 현대사회에서 유병률이 꾸준히 증가하고 있으며, 삶의 질을 크게 떨어뜨리는 주요 요인으로 작용한다(Cacciatore 등, 2025; Kim 등, 2018; Lee 등, 2022; Tang 등, 2025). 최근에는 천연물과 발효 유래 성분이 면역 균형을 회복하고 염증 반응을 완화하며, 알레르기의 치료 및 예방에 효과가 있다는 가능성을 제시한 보고가 나오고 있어 약물치료의 대안으로 주목받고 있다(Dębińska와 Sozańska, 2022; Li 등, 2022; Zhu 등, 2022).
이러한 기존 연구들을 종합적으로 봤을 때 유산균 발효 기반 효능 연구는 기능성 식품 산업뿐 아니라 공중보건적 측면에서도 중요한 의미를 지닌다고 판단된다. 현재 다양한 천연물 기반 소재에 대한 연구는 활발하나, 시중의 대부분의 유산균 소재들은 기개발된 유산균을 단순 혼합하여 제조한 원료성 제품들이 주를 이루고 있으며 본 논문에서의 내용과 같이 유산균 발효를 통한 천연물 소재 연구는 미흡한 실정이다.
따라서 본 연구는 느타리버섯을 P. pentosaceus로 발효하고 그 추출물로서 항염증 활성을 검증함으로써 다양한 소재 개발에 응용할 수 있는 기초자료를 확보함과 동시에 식품의 기능성과 안전성을 확보하여 국민 생활 증진 및 관련 농가 소득 향상에 기여하고자 하였다.
2. 재료 및 방법
실험에 사용한 Pleurotus ostreatus는 장흥군버섯산업연구원(Jangheung, Korea)으로부터 제공받아 사용하였다.
본 실험에 사용한 세포배양 관련 시약인 Streptomycin, Penicillin, Fetal Bovine Serum (FBS)과 Dulbecco’s modified Eagle’s medium (DMEM)는 Invitrogen (Carlsbad, CA, USA)에서 구입하였으며 분석 실험에 사용된 시약과 유기용매(Sigma-Aldrich Co., St. Louis, MO, USA)는 제품 중 일급 이상 등급을 선택하여 사용하였다.
실험에 사용된 유산균은 장흥군 및 임실군에서 수집하고 임실치즈앤식품연구소에서 분리 동정한 균주 Lactobacillus plantarum ICFPL-001, Leuconostoc lactis ICFPL-004, L. sakei LJ001, L. sakei LI033, L. sakei LGy039, Pediococcus pentosaceus ALJ015, P. pentosaceus ALJ024이며, 또한 동일한 방법으로 동정된 L. acidophilus KCTC18859P 및 P. pentosaceus KCTC18860P 균주는 본 연구진이 생물자원센터(KCTC)에 기탁한 특허 균주를 사용하였다. 각각의 유산균 배양은 MRS broth (Difco, Sparks, MD, USA)를 이용하여 지정된 조건에서 배양하였다.
건조된 느타리버섯(P. ostreatus) 자실체를 분말화한 후, 건조중량 대비 10배(w/v)의 증류수를 가하여 80°C에서 4시간 동안 열수추출하였다. 추출액은 8 μm 여과지를 사용하여 진공 여과하였으며, 여과액을 동결건조하여 분말 형태의 느타리버섯 추출물을 제조하였다. 동결건조 추출물의 수율을 확인하였으며, 제조된 추출물은 사용 전까지 -20°C에서 보관하였다.
유산균 발효를 위하여 동결건조 느타리버섯 추출물을 증류수에 재용해하여, 최종 발효액 기준 느타리버섯 건조중량 환산 농도가 5%(w/v)가 되도록 조성하였다. 이때 추출물 용액은 전체 발효액 부피의 50%(v/v)를 차지하도록 하였다. 유산균의 안정적인 생육과 발효 중 유기산 생성 촉진을 위하여 sucrose 2%(w/v)와 lactose 3%(w/v)를 탄소원으로 첨가하였다. 조성된 발효 전 추출물은 autoclave를 이용하여 121°C, 1.2 atm 조건에서 15분간 멸균한 후, 무균상 내에서 실온까지 방랭하였다.
유산균 접종을 위해 사용한 균주는 MRS broth (Difco, Sparks, MD, USA)에서 37°C, 18시간 동안 전배양한 후, van Bokhorst-van de Veen 등(2012)의 방법에 따라 대수증식기에 해당하는 균체 현탁액(OD600=1.0)을 사용하였다. 전배양된 유산균 배양액을 멸균된 발효배지에 최종 발효액 대비 1%(v/v)가 되도록 접종하였으며, 37°C로 설정된 배양기에서 48시간 동안 정치 배양하였다. 발효 종료 후 시료를 회수하여 이후 이화학적 및 생리활성 분석에 사용하였다.
총산도는 시료를 원심분리한 후 상등액 10 mL를 취하여 분석하였다. 시료는 1% phenolphthalein 용액을 지시약으로 첨가하고, 0.1 N NaOH 표준용액으로 중화될 때까지 적정하였다. 적정 결과는 acetic acid 함량으로 환산하여 총산도(%)로 나타내었으며, 젖산의 경우 0.1 N NaOH 1 mL가 젖산 0.009 g에 해당하는 것으로 계산하여 백분율(%)로 표현하였다.
총폴리페놀 함량은 Folin-Denis법을 이용하여 측정하였다(Singleton과 Rossi, 1965). Ethanol을 사용해 1 mg/mL 농도로 희석한 시료와 Folin-Denis reagent를 각 100 μL씩 혼합한 뒤, 실온에서 3분간 반응시켰다. 이후 10% sodium carbonate 용액 100 μL를 첨가한 뒤 잘 혼합하였으며 1시간 동안 반응을 진행하였다. 반응이 끝난 후 상등액을 분리하여 microplate reader (Molecular Devices, Sunnyvale, CA, USA)를 사용해 760 nm 파장에서 흡광도를 측정하였다. 정량 분석은 gallic acid를 표준물질로 하여 작성한 표준곡선을 기준으로 수행하였다.
총플라보노이드 함량 측정은 Davis (1947)의 측정법을 이용하였다. 시료 100 μL (1 mg/mL, ethanol 희석)을 취해 diethylene glycol 1 mL를 넣고 혼합한 뒤, 1 N NaOH 용액 100 μL를 추가하여 균질화하였다. 이후 혼합액을 37°C water bath에서 1시간 동안 반응시킨 후, microplate reader (Molecular Devices, Sunnyvale, CA, USA)를 이용해 420 nm 파장에서 흡광도를 측정하였다. 총플라보노이드 함량은 naringin을 표준물질로 하여 작성한 표준곡선을 바탕으로 산출하였다.
세포 생존율 변화를 관찰하기 위해 RAW 264.7 세포, RBL-2H3 세포, AGS 세포를 사용하였다. 각 세포는 10% FBS, 1X antibiotic-antimycotic이 첨가된 DMEM 배지에서 37°C, CO2 5% 조건하에 배양하였다. 세포가 70-80% 수준으로 증식하면 Trypsin-EDTA로 처리하여 계대배양을 진행한 세포를 사용하였다.
세포 생존율은 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) 환원법을 사용하여 측정하였다. 세포를 96-well plate에 분주한 다음 12시간 동안 배양하였으며, 이후 농도별 시료를 처리하고 추가로 24시간 동안 배양하였다. 이후 MTT 용액(0.5 mg/mL)을 첨가하고 37°C에서 4시간 배양하고 MTT를 환원시켜 생성된 formazan이 배지에서 분리되지 않도록 배지를 제거하였다. DMSO를 100 μL 가하여 10분동안 혼합한 후 540 nm에서 흡광도를 측정하였다.
Nitric oxide (NO) 생성량은 griess 시약을 이용하여 측정하였다. 세포를 24-well plate에 분주한 후 12시간 동안 배양하고, 농도별로 시료를 처리하였다. 이후 Escherichia coli O55:B5 유래 lipopolysaccharide (LPS, 1 μg/mL)를 첨가하여 20시간 추가 배양하였다. 배양 후 세포 배양액 100 μL와 100 μL의 griess 시약(1% sulfanilamide in 5% phosphoric acid 및 1% α-naphthylamide in H2O)을 혼합하여 실온에서 10분간 반응시킨 뒤, microplate reader (Molecular Devices, Sunnyvale, CA, USA)를 이용해 550 nm 파장에서 흡광도를 측정하였다. sodium nitrite (NaNO2)를 이용하여 표준곡선을 작성하여 NO 생성량을 산출하였다. 배지만 처리한 세포를 음성 대조군으로 하고 LPS만 처리한 세포를 양성 대조군으로 설정하여 시료 처리군과 비교하였다.
세포를 농도별 추출물로 4시간 동안 전처리한 후, 1 μg/mL LPS에 24시간 동안 노출하였다. 배양 후 상층액을 수거하여 TNF-α, PGE2, IL-1β 수치를 측정하기 위하여 ELISA를 수행하였다. 코팅되어 있는 96 well plate에 각 분석 대상 사이토카인의 primary antibody를 100 μL씩 넣고 4°C에서 overnight 반응시켰다. 그 후 0.5% tween 20 wash 용액을 사용하여 5분씩 3회 wash한 후 샘플과 standard 용액을 100 μL씩 넣고 2시간 동안 상온에서 반응시켰다. 동일한 방식으로 wash한 다음 HRP- conjugated secondary antibody를 각 100 μL씩 첨가하여 2시간 동안 상온에서 반응시킨 후 다시 wash한 후 avidin/biotin을 가하여 발색시켜 흡광도를 측정하였고, standard의 흡광도로부터 표준곡선을 구해 이를 활용하여 각 샘플의 흡광도값을 기반으로 사이토카인의 양을 정량하였다.
탈과립은 Matsuda 등(2002)의 β-hexosaminidase 측정법을 이용하여 확인하였다. Dinitrophenyl-immunoglobulin E (DNP-IgE)가 포함된 DMEM 배지를 이용하여 세포를 24-well plate에 분주하고 배양하였다. 이후 세포를 siraganian buffer (119 mM NaCl, 5 mM KCl, 5.6 mM glucose, 0.4 mM MgCl2, 25 mM PIPES, 1 mM CaCl2, 0.1% BSA, pH 7.2)로 세척한 뒤, 동일한 buffer에서 37°C 조건으로 10분간 전배양하였다. 다음으로, 시료를 농도별로 희석하여 세포에 처리하고 30분간 반응시킨 후 DNP-BSA를 첨가하여 2시간 동안 반응하였다. 반응 후 ice bath에서 10분간 냉각하여 반응을 종료시켰다. 상층액 40 μL에 substrate buffer (2 mM 4-p-nitrophenyl-N-acetyl-β-D- glucosaminide, 0.05 M sodium citrate, pH 4.5) 40 μL를 첨가하여 37°C의 조건에서 1시간 동안 반응시킨 후, stop solution (0.1 M Na2CO3/NaHCO3, pH 10.5) 200 μL를 첨가하여 반응을 중지하였다. 이후 흡광도는 405 nm에서 측정하였다.
Histamine은 Shore 등(1959)의 방법으로 진행하였다. 세포 배양액 25 μL에 0.1 N HCl 22.5 μL와 60% HClO4 42.5 μL를 첨가하여 반응시킨 뒤 원심분리하였다. 분리된 상층액 40 μL에 5 N NaOH 25 μL, n-butanol 500 μL, NaCl 0.06 g을 넣고 혼합하여 진탕한 후 다시 원심분리하였다. 이어서 butanol층 400 μL를 취해 0.1 N HCl 150 μL, n-heptane 0.5 mL를 혼합하고 진탕 후 원심분리하였다. 분리된 water층 100 μL에 1 N NaOH 200 μL와 0.1% o-phthaldialdehyde 용액 5 μL를 가하여 37°C에서 3분간 반응시켰으며, 이어 3 N HCl 10 μL를 첨가해 2분간 반응을 지속하였다. 반응이 완료된 시료의 흡광도는 360 nm (excitation)와 450 nm (emission)에서 측정하였다. 대조군은 음성 대조로 배지만 처리한 세포, 양성 대조로 LPS 또는 DNP-BSA만 처리한 세포를 설정하여 시료 처리군의 활성을 비교하였다.
3. 결과 및 고찰
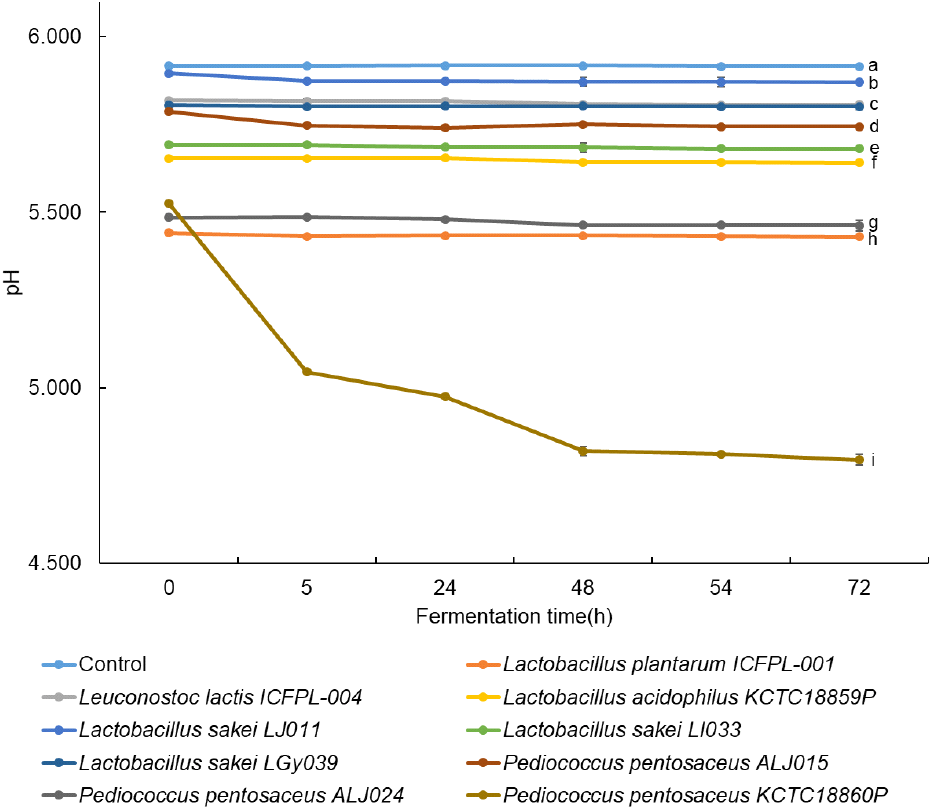
L. plantarum ICFPL-001, Leu. lactis ICFPL-004, L. acidophilus KCTC18859P, L. sakei LJ001, L. sakei LI033, L. sakei LGy039, P. pentosaceus ALJ015, P. pentosaceus ALJ024, P. pentosaceus KCTC18860P를 각각 접종한 느타리버섯 발효물의 발효기간 동안 pH를 측정한 결과는 Fig. 1과 같다. 발효가 진행됨에 따라 P. pentosaceus KCTC18860P를 접종한 발효물에서 pH가 5.525에서 4.794로 가장 뚜렷하게 감소하는 것을 확인할 수 있었다. 한편, 본 연구에서 사용된 비발효 대조군(control)은 발효 종료 시점의 발효물과 동일한 pH로 보정한 비발효 느타리버섯 추출물로 설정하여, 관찰된 생리활성 변화가 단순한 pH 감소에 기인하는지 여부를 구분하기 위한 대조군으로 사용하였다. Xu 등(2023)은 염장 생선에서 분리한 P. pentosaceus 균주가 빠른 유기산 생성으로 pH가 효과적으로 감소한다고 보고하였으며, 본 연구에서도 유사한 경향을 보였다.
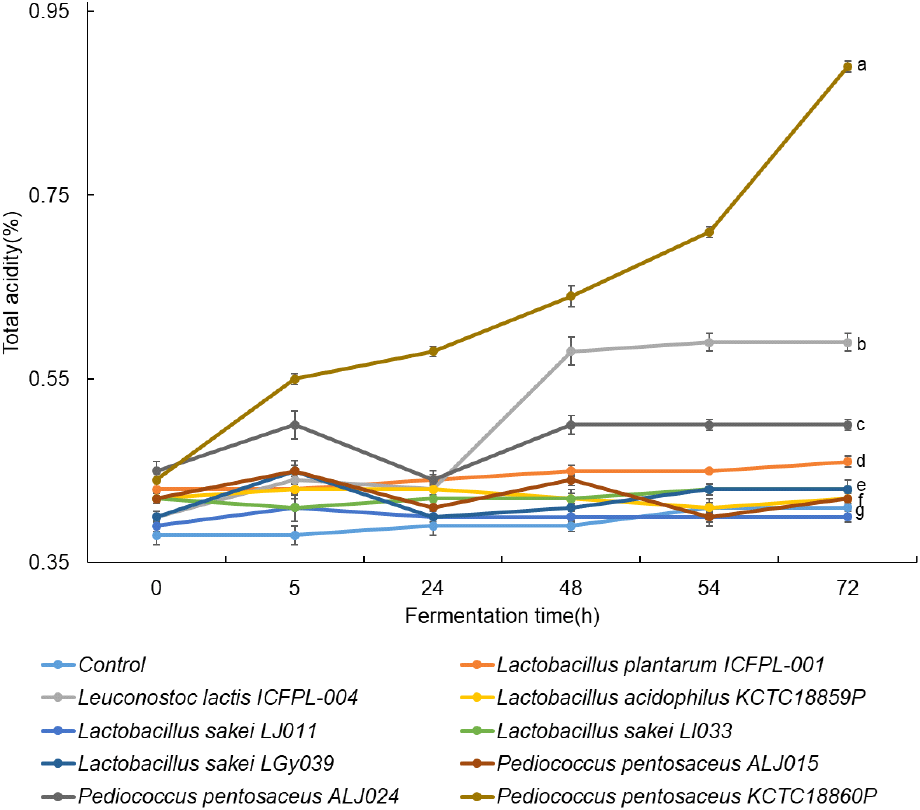
L. plantarum ICFPL-001, Leu. lactis ICFPL-004, L. acidophilus KCTC18859P, L. sakei LJ001, L. sakei LI033, L. sakei LGy039, P. pentosaceus ALJ015, P. pentosaceus ALJ024, P. pentosaceus KCTC18860P를 각각 접종한 느타리버섯 발효물의 발효기간 동안 총산도를 측정한 결과는 Fig. 2와 같다. 발효가 진행됨에 따라 P. pentosaceus KCTC18860P를 접종한 발효물에서 총산도가 0.44에서 0.89로 가장 크게 증가하였다. Meng 등(2025)은 P. pentosaceus는 당을 젖산으로 효율적으로 전환하여 총산도를 크게 높이는 것으로 알려져 있다고 보고하였고, Nielsen(2017)은 시료 내 산의 축적이 많아질수록 적정에 필요한 염기량이 증가하고, 이에 따라 총산도가 높게 측정되며, 동시에 자유 수소이온 농도 증가로 인해 pH가 낮아지게 된다고 보고하였다. 특히 본 연구에서는 발효 종료 시점의 발효물과 동일한 pH로 보정한 비발효 느타리버섯 추출물을 대조군으로 설정하였음에도 불구하고, 총산도는 발효 균주에 따라 유의한 차이를 보였다. 이는 동일한 pH 조건 하에서도 발효 여부 및 접종 균주에 따라 생성 ․ 축적되는 유기산의 총량과 특성이 상이함을 시사하며, 총산도는 단순한 산성화 효과와 발효 과정에서의 대사적 변화를 구분하는 지표로 해석될 수 있다. 따라서 본 연구에서는 pH를 낮추고 산도를 높여 유산균 발효 활성이 우수하게 나타난 P. pentosaceus KCTC18860P를 최종 발효 균주로 선정하였다.
폴리페놀 화합물은 항산화, 항염증, 항암, 신경보호 등 다면적인 생리활성 기능을 가진다고 알려져 있으며 플라보노이드는 식물 유래의 주요 폴리페놀 계열 화합물로, 항산화 활성뿐 아니라 다양한 생리활성 기능을 가진 것으로 알려져 있다(Hedayati 등, 2023; Heim 등, 2002).
느타리버섯 농축액과 이를 P. pentosaceus로 발효한 느타리버섯 발효물 총 2종에 대해 총폴리페놀 및 총플라보노이드 함량을 측정한 결과는 Table 1과 같다. 총폴리페놀 함량은 느타리버섯 농축액 20.85 mg/g, P. pentosaceus 느타리버섯 발효물 11.55 mg/g으로 나타났다. 이러한 결과는 모든 유산균 발효에서 폴리페놀 함량이 증가하는 것은 아니며, 다양한 조건의 상호작용에 따라 정체 또는 감소가 관찰될 수 있고 실제로 꽃송이버섯에서는 유산균 발효가 폴리페놀 함량을 크게 변화시키지 않았다는 보고와 유사하다고 볼 수 있다(Lee 등, 2016). 유산균에 의해 분해, 변형되어 생성된 새로운 대사체, 유기산 및 미세 성분이 항산화 및 항염증 효과를 갖는 경우가 보고되었으며 총폴리페놀 함량 감소가 반드시 항염증 및 항알레르기 효과 감소로 이어지지 않고, 오히려 발효로 생성된 다른 생리활성 성분들이 기능성을 증진할 수 있음을 고려할 필요가 있다(Grondalska와 Kolniak-Ostek, 2025; Yang 등, 2023; Zhang 등, 2025).
| Samples | Content (mg/g) | |
|---|---|---|
| Total polyphenol | Total flavonoid | |
| PE1) | 20.85±0.202)*3) | 1.85±0.08 |
| PPPE | 11.55±0.18 | 2.02±0.11 |
총플라보노이드 함량은 P. pentosaceus 느타리버섯 발효물 2.02 mg/g, 느타리버섯 농축액은 1.85 mg/g 순으로 나타났다. 이러한 결과와 유사하게 유산균 발효가 꽃송이버섯의 총플라보노이드와 항산화능을 동반 향상시키는 경향은 논문으로도 보고되고 있다(Lee 등, 2016). 추가적으로, 본 연구에서 느타리버섯 발효물의 총폴리페놀 함량이 낮아졌음에도 불구하고 유산균 발효물이 잠재적인 생리활성을 지닐 수 있음은 여러 선행 연구에서도 관찰된다. 느타리버섯 발효물의 항염, 항알레르기 활성은 단순 총폴리페놀 및 플라보노이드 증가만으로 특정하기 어렵고, 문헌에서는 버섯류의 inotodiol, ergosterol 및 유도체, Ganoderma계 ganoderic acids, β-glucan 및 다당류 등이 비만세포 탈과립 억제를 통한 항알레르기 효과와 항염 효과를 나타내는 것으로 보고되어 있다(Kageyama-Yahara 등, 2010; Kohda 등, 1985; Ma 등, 2013; Nguyen 등, 2020; Sheng 등, 2020; Shi 등, 2025; Won 등, 2023). 따라서 이러한 결과를 바탕으로 버섯을 유산균 발효함으로서 증가한 플라보노이드를 통해 다양한 생리활성 기능의 향상 가능성이 있을 것으로 사료된다.
세포 생존율은 독성 평가의 대표적이며 표준적인 지표이며, 이를 통해 세포독성을 확인할 수 있다(Gatto 등, 2023; Jhamb 등, 2025). 각 동물 세포주들을 96 well에서 배양하여 느타리버섯 추출물과 발효물을 0 μg/mL, 50 μg/mL, 100 μg/mL, 300 μg/mL, 500 μg/mL, 700 μg/mL, 1000 μg/mL 농도로 24시간 동안 처리하여 배양한 후 MTT assay를 수행하였다. 각 추출물의 세포 생존율을 평가한 결과는 Fig. 3과 같다.
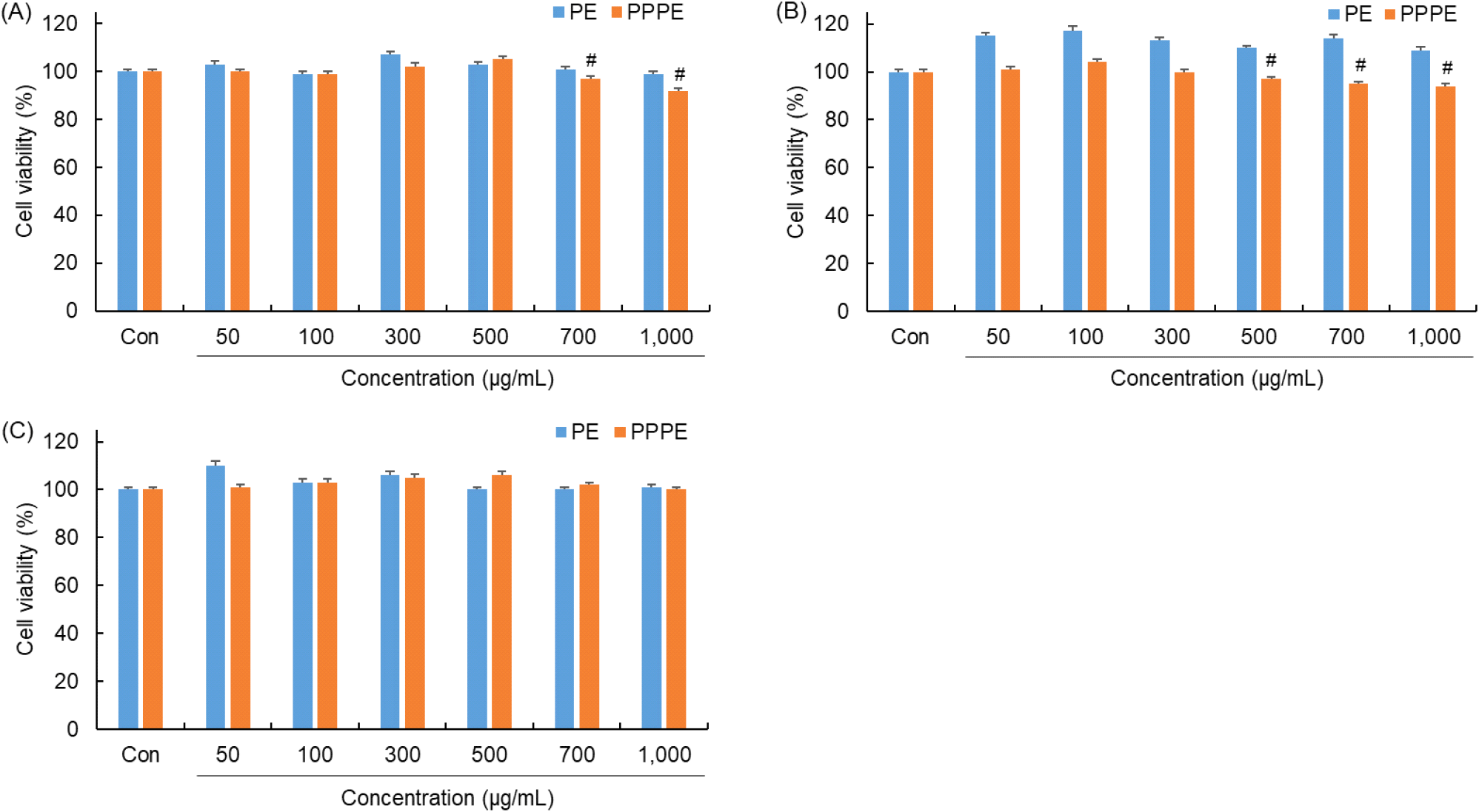
RAW 264.7 세포, RBL-2H3 세포, AGS 세포에 P. pentosaceus 느타리버섯 발효물과 느타리 추출액을 처리한 후 확인한 결과, 세포 생존율이 감소하지 않았음을 확인하였다. 즉, 이들 시료에서는 유의한 세포독성이 관찰되지 않은 것으로 판단된다. Jedinak 등(2011)에 따르면 느타리버섯은 세포독성이 없다고 보고하고 있다. 더불어, Choi 등(2021)에 따르면 P. pentosaceus와 L. acidophilus로 발효한 표고버섯은 세포독성이 없다고 보고한 결과와 유사한 경향을 확인할 수 있다. 따라서 느타리버섯 유산균 발효는 세포 생존율에 영향을 끼치지 않는 것으로 판단된다.
일산화질소(nitric oxide, NO)는 면역반응에 중요한 역할을 하지만, 과도하게 생성될 경우 염증 매개 물질로 작용하여 조직 손상을 비롯한 다양한 염증성 질환을 유발할 수 있다(Bogdan, 2001). NO 생성에 대한 느타리버섯 추출물과 발효물의 억제 효과를 측정하였다. 느타리버섯 추출물과 발효물을 0 μg/mL, 300 μg/mL, 400 μg/mL, 500 μg/mL 농도로 4시간 동안 전처리하여 LPS를 처리하고 Griess 시약을 이용하여 세포 배양액에 존재하는 생성된 NO 양을 분석한 결과는 Fig. 4와 같다.
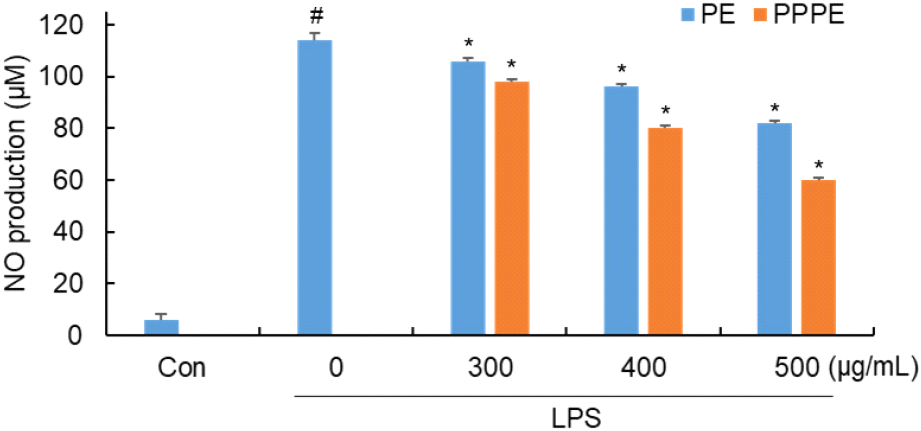
LPS만 단독 처리한 군에서는 114 μM의 NO가 생성되었으며 느타리 추출물의 경우에는 106 μM, 96 μM, 82 μM, P. pentosaceus 느타리버섯 발효물의 경우에는 98 μM, 80 μM, 60 μM의 NO 생성을 나타내었다. 이러한 결과는 각 추출물이 LPS에 의해 세포 내에 유도된 NO의 생성을 저해하는 것으로 확인하였으며, 발효물의 저해 효과가 더 높게 나타난 것을 확인할 수 있다. Gunawardena 등(2014)에 따르면 느타리버섯이 NO 생성을 억제한다고 보고하고 있다. 더불어, Choi 등(2021)에 따르면 P. pentosaceus와 L. acidophilus로 발효한 표고버섯은 세포독성 없이, NO 생성을 억제한다고 보고하고 있다. 따라서 본 실험을 통한 유산균 발효는 세포독성 우려 없이 NO 생성억제 효과를 높일 수 있는 것으로 사료된다.
사이토카인은 염증반응을 매개하는 주요 인자로, 대식세포 등 면역 관련 세포에서 분비되어 다양한 염증 반응을 유도한다. 이 가운데 tumor necrosis factor (TNF)-α와 interleukin (IL)-1β는 대표적인 염증성 사이토카인으로 알려져 있으며, TNF-α는 세포 증식, 생존, 분화 및 사멸을 조절할 뿐 아니라 염증 반응 시 발현이 증가하여 IL-1β와 같은 염증성 사이토카인의 생성을 촉진하는 주도적 역할을 수행한다. IL-1β는 면역 방어와 조직 손상에 대한 반응에 관여하며, prostaglandin E2 (PGE2)의 생성을 유도함으로써 염증 반응을 증폭시킨다(Bonaventura 등, 2018; Bradley, 2008; Dinarello, 2009; Kany 등, 2019; Neeb 등, 2011; Rieser 등, 1997). 느타리버섯 추출물과 발효물을 0 μg/mL, 100 μg/mL, 200 μg/mL, 300 μg/mL 농도로 4시간 동안 전처리한 후 LPS를 처리한 후 ELISA를 이용하여 RAW 264.7 세포 배양액 중에 존재하는 IL-1β, TNF-α, PGE2의 양을 분석한 결과는 Fig. 5와 같다.
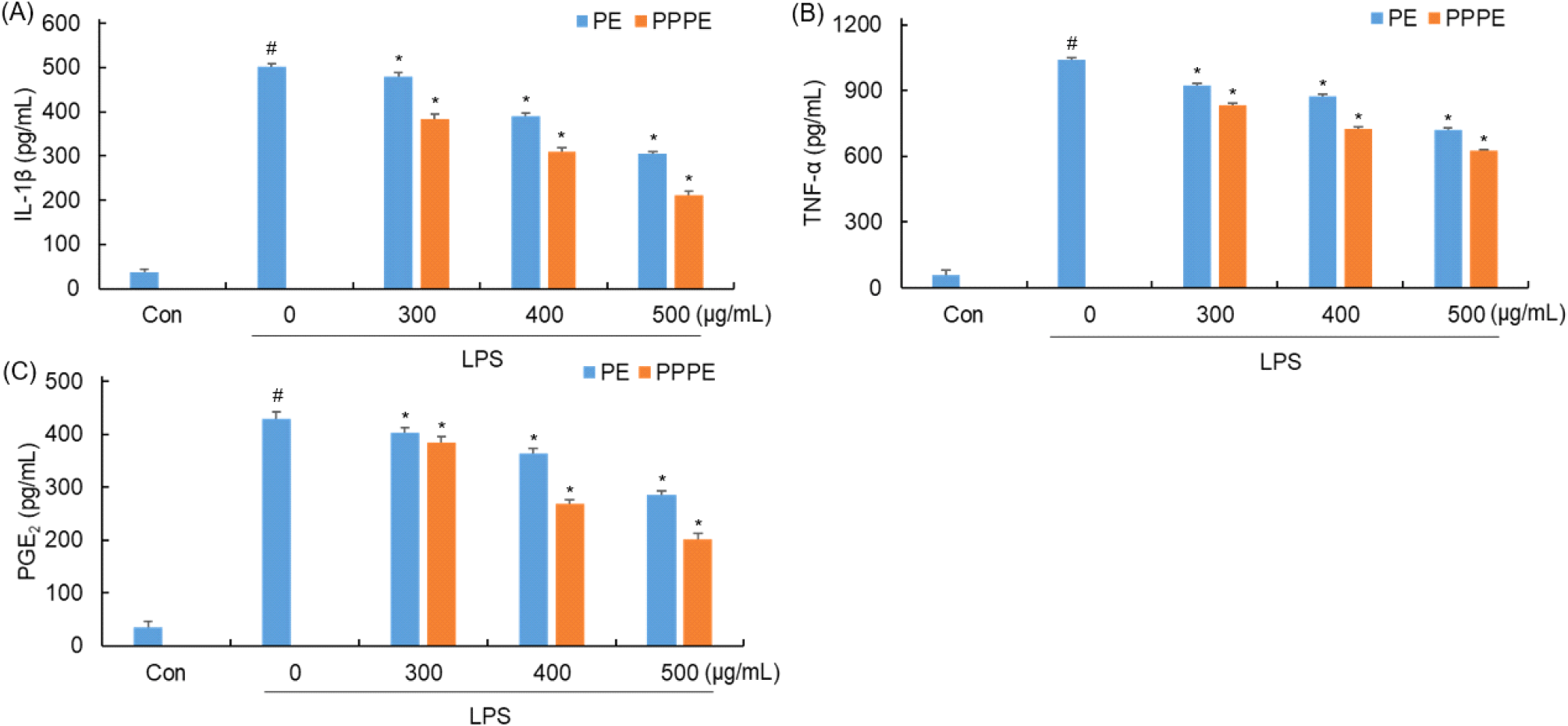
LPS만 단독 처리한 군에서는 502 pg/mL의 IL-1β가 생성되었으며 느타리 추출물의 경우에는 480 pg/mL, 390 pg/mL, 305 pg/mL, P. pentosaceus 느타리버섯 발효물의 경우에는 383 pg/mL, 309 pg/mL, 211 pg/mL의 IL-1β의 생성량을 나타내었다.
LPS만 단독 처리한 군에서는 1039 pg/mL의 TNF-α가 생성되었으며 느타리 추출물의 경우에는 923 pg/mL, 873 pg/mL, 720 pg/mL, P. pentosaceus 느타리버섯 발효물의 경우에는 834 pg/mL, 727 pg/mL, 624 pg/mL의 TNF-α 생성을 나타내었다.
LPS만 단독 처리한 군에서는 430 pg/mL의 PGE2가 생성되었으며 느타리 추출물의 경우에는 404 pg/mL, 363 pg/mL, 286 pg/mL, P. pentosaceus 발효 느타리 추출물의 경우에는 384 pg/mL, 268 pg/mL, 201 pg/mL의 PGE2 생성을 나타내었다. 발효물과 비발효물의 cytokine 생성 억제 효과를 보면 IL-1β, TNF-α, PGE2 모두 농도 의존적으로 발현량이 감소하는 경향을 보였으며 비발효물보다 발효물에서 더 큰 억제 효과를 나타내었다. Jedinak 등(2011)에 따르면 느타리버섯의 TNF-α, IL-6, IL-12p40, PGE2 억제능을 보고하고 있다. 더불어, Choi 등(2021)에 따르면 세포독성 없이 P. pentosaceus와 L. acidophilus로 발효한 표고버섯 발효물이 비발효 대비 IL-1β, IL-4, TNF-α, PGE2를 억제한다고 보고한 결과와 유사한 경향을 확인할 수 있다. 따라서 유산균 발효물이 비발효물 대비 cytokine 생성을 억제하는 효능을 가지고 있는 것으로 판단된다. 발효 과정에서 생성된 유기산, 단쇄지방산 등이 대식세포의 NF-κB 경로를 억제함으로써 NO 및 염증성 cytokine 생성을 감소시킬 수 있음이 보고되고 있다(Park 등, 2005; Yang, 2020). 본 연구에서는 이러한 분자적 기전을 직접 확인하지는 않았으나, 기존 문헌과 유사한 농도 의존적 억제 패턴을 확인함으로서 발효물의 항염증 잠재력을 시사할 수 있다. 향후 NF-κB 활성 분석 등 분자 수준 검증 연구가 필요하다.
β-Hexosaminidase는 비만세포 탈과립 시 히스타민과 함께 방출되는 대표적인 가수분해 효소로, 알레르기 반응에서 비만세포 활성화의 지표로 활용된다(Moon 등, 2014). 한편, 히스타민(histamine)은 비만세포 탈과립 시 탈과립 과정에서 분비되는 대표적인 염증 매개물질로, 혈관 확장, 점액 생성 증가, 기관지 수축 등을 유발하여 알레르기 반응의 주요 증상을 매개한다(Galli와 Tsai, 2012). 느타리버섯 추출물과 발효물을 0 μg/mL, 300 μg/mL, 400 μg/mL, 500 μg/mL 농도로 처리 한 후 탈과립화 및 histamine 생성 억제 효과를 확인하기 위해 RBL-2H3 세포 배양액에 존재하는 β-hexosaminidase를 분석한 결과는 Fig. 6과 같다.
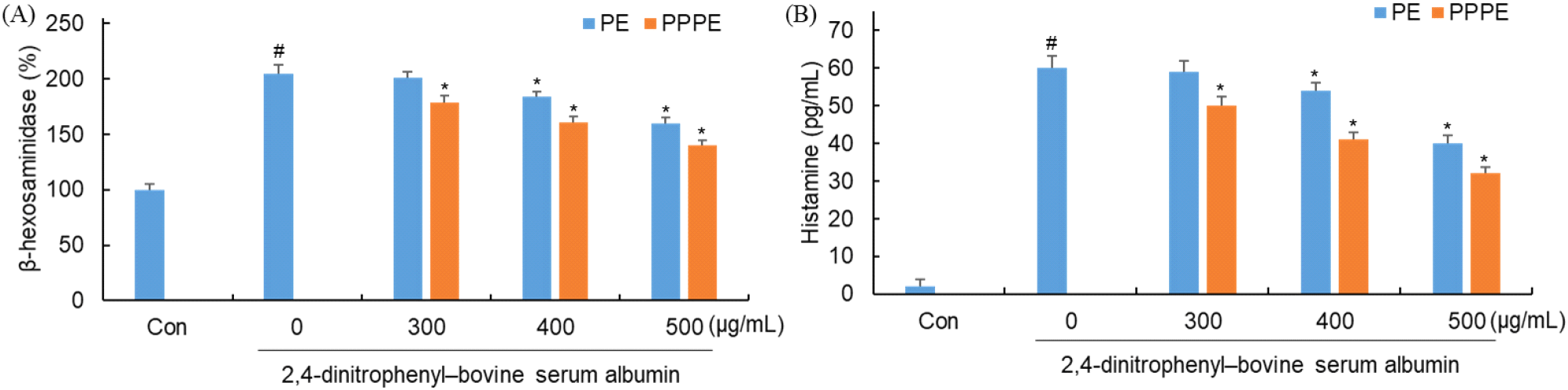
DNP-IgE와 DNP-BSA만 단독 처리한 군에서는 205%의 β- hexosaminidase 활성을 나타냈으며 느타리 추출물의 경우에는 201%, 184%, 160%, P. pentosaceus 느타리버섯 발효물의 경우에는 179%, 161%, 140%의 β-hexosaminidase 활성을 나타내었다.
DNP-IgE와 DNP-BSA만 단독 처리한 군에서는 60 pg/mL으로 histamine 생성이 감소하였으며 느타리 추출물의 경우에는 59 pg/mL, 54 pg/mL, 40 pg/mL, P. pentosaceus 느타리버섯 발효물의 경우에는 50 pg/mL, 41 pg/mL, 32 pg/mL의 histamine 생성을 나타내었다. 2종의 탈과립화 및 histamine생성 억제 효과를 보면 모두 농도 의존적으로 활성이 감소하는 경향을 보였으며 비발효물보다 발효물에서 더 큰 억제 효과를 나타내었다. 따라서 이러한 결과를 토대로 느타리버섯 발효물을 활용한다면 식품의 과민반응 완화 보조 가능성을 지니고 있다고 판단된다. Yan과 Choi(2019)에 따르면 표고버섯 메탄올 추출물이 농도 의존적으로 탈과립을 억제하고 histamine 생성을 감소시킨다고 보고하고 있다. Lee 등(2019)에 따르면 표고, 느타리, 팽이 물 추출물의 β-hexosaminidase 억제 활성을 보고하고 있다. Phull 등(2021)에 따르면 동충하초를 유산균 발효시킨 추출물이 비발효 대비 탈과립을 더 크게 억제한다고 보고한 결과들과 유사한 경향을 확인할 수 있었다. 이러한 결과를 통해 느타리버섯 발효물은 β-hexosaminidase 및 histamine 분비를 억제함으로써 알레르기 증상을 개선 효과가 있을 것으로 판단된다.
4. 요약
본 연구에서는 P. pentosaceus 발효를 통해 느타리버섯의 항염증 활성을 향상시키는 것을 목적으로 수행되었다. 다양한 유산균 중 적절한 유산균을 선별하는 과정에서 뚜렷하게 pH가 낮아지고 총산도가 높아지는 P. pentosaceus를 최종 균주로 선발하고 P. pentosaceus를 이용하여 느타리버섯 발효물을 제조하여 비발효물과의 비교 분석한 결과, 총폴리페놀 함량은 발효물이 비발효물에 비해 낮지만 총플라보노이드 함량은 높은 것을 확인하였으며 RAW 264.7 세포, RBL-2H3 세포, AGS 세포 처리에 의한 세포 생존율에는 영향이 없는 것을 확인하였다. 또한, 비발효물보다 발효물에서 NO 생성 억제 효과가 높아졌으며 염증성 사이토카인 IL-1β, TNF-α, PGE2의 생성이 모두 유산균 발효물 처리에 의해서 감소되었음을 확인하였다. 느타리버섯 추출물만을 처리한 군보다 유산균 발효를 적용한 추출물에서 β-hexosaminidase와 histamine의 분비량이 감소함을 확인하였다. 이러한 결과는 in vitro 기반의 기초 연구로서, 느타리버섯 P. pentosaceus 발효물이 항염증 및 항알레르기 기능성 식품 소재로 활용될 가능성을 시사하며, 향후 동물실험 및 인체적용시험을 통한 추가 검증이 필요할 것으로 판단된다.