








1. 서론
한국이 급격한 초고령화 사회로 진입함에 따라, 노화에 기인한 근감소증(sarcopenia)이 중요한 문제로 대두되고 있다. 국가통계포털(KOSIS)에 따르면, 2025년 기준 전체 인구의 약 20%가 65세 이상에 해당할 정도로 고령인구가 급증하였으며, 이러한 인구 구조 변화는 보건의료 및 복지 체계 전반에 큰 영향을 미치고 있다(KOSIS, 2025). 고령 인구는 다양한 만성질환의 발병 위험이 높으며, 그 중에서도 근감소증은 2021년에 질병코드가 부여된 중요한 노인성 질환으로 주목받고 있다. 근감소증은 근육량, 근력 및 신체 기능이 감소되어 발생하는 질환으로 정의되며, 주로 65세 이상에서 유병률이 높아진다(Chen 등, 2020; Cruz-Jentoft 등, 2019). 이러한 근감소증으로 인한 신체 활동의 제약은 노인의 자립적 생활 능력을 저하시킬 뿐만 아니라 낙상 및 골절의 위험을 높여 입원 및 장기요양서비스 이용률을 증가시키는 등 사회 ․ 경제적 부담을 가중시킨다(Antunes 등, 2017; Bruyère 등, 2019; Veronese 등, 2019). 또한 당뇨병, 비만, 심혈관 질환 등 만성 질환의 유병률을 높여, 단순한 노화 현상을 넘어 공중보건 측면에서도 중요한 관리 대상 질환으로 인식되고 있다(Beaudart 등, 2014; Beaudart 등, 2025). 따라서 근감소증의 예방과 관리를 위한 전략 마련은 초고령사회에서 건강수명을 연장하고 의료비 부담을 완화하기 위한 필수 과제로 고려되고 있다.
근감소증의 발생은 산화 스트레스(oxidative stress)의 증가, 단백질 합성과 분해의 불균형, 만성 염증 등이 복합적으로 작용하는 것으로 알려져 있다(Kim 등, 2023b). 노화가 진행됨에 따라 활성산소종(reactive oxygen species, ROS)의 축적으로 산화 스트레스가 증가하며, 이는 근육 내 염증을 증가시켜 단백질 항상성(protein homeostasis) 불균형을 초래한다(Kim 등, 2023a; López-Otín 등, 2023). 단백질 항상성은 세포 내 단백질의 합성, 접힘(folding), 분해 과정이 정교하게 조절되는 생리적 균형 상태를 의미하며, 정상적인 근육세포에서는 단백질의 합성과 분해가 균형을 이루어 세포 내 단백질 항상성이 유지된다(Tan 등, 2020). 그러나 노화나 스트레스와 같은 외부 자극으로 인해 이 균형이 무너질 경우, 단백질 변성 및 손상이 누적되어 근섬유 단백질의 손실로 이어진다(Larsson 등, 2019). 이러한 근육 세포의 단백질 항상성 불균형의 유도를 위해, 합성 글루코코르티코이드(glucocorticoid)인 덱사메타손(dexamethasone, Dex)이 널리 사용된다(Wang 등, 2023).
Dex는 글루코코르티코이드 수용체(glucocorticoid receptor, GR)를 통해 근감소증을 유도하는 약물이다. Dex는 GR에 결합하여 이를 활성화하며, Regulated in development and DNA damage response 1 (Redd1)과 Krüppel-like factor 15 (Klf15)의 발현이 유도되어 단백질 합성 경로는 억제되고 분해 경로는 촉진되어 근위축이 발생한다(Tsuchida 등, 2017). Redd1에 의한 단백질 합성 경로의 억제는, Redd1이 tuberous sclerosis (TSC)1/2 복합체를 활성화하고 ras homolog enriched in brain (Rheb)을 억제하여 mammalian target of rapamycin complex 1 (mTORC1) 활성을 직접적으로 저해함과 동시에, protein phosphatase 2A (PP2A)를 활성화하고 protein kinase B (Akt)를 탈인산화함으로써 불활성화시켜 Akt의 하위 기전인 mTORC1 신호 전달을 간접적으로 감소시키는 이중 기전을 통해 일어난다(Dennis 등, 2014). Klf15에 의한 단백질 분해 경로의 촉진은, Klf15가 forkhead box O3α (FoxO3α)의 발현을 높여 유비퀴틴-프로테아좀 시스템(ubiquitin-proteasome system)의 핵심 효소이며 E3 ubiquitin ligase인 Muscle RING-finger protein-1 (MuRF1)과 muscle atrophy F-box (Atrogin-1)의 전사를 촉진함으로써 일어난다(Permpoon 등, 2025). 이러한 변화는 근육 내 단백질 항상성을 손상시켜 근육 단백질 손실을 촉진하며, 결과적으로 근섬유 위축 및 근력 감소를 초래한다(Braun과 Marks, 2015; Lee 등, 2021). 또한, Dex 처리로 인한 ROS의 축적과 항산화 방어체계(superoxide dismutase, SOD; catalase, CAT; glutathione peroxidase, GPx 등의 항산화 효소)의 기능 저하는 산화 스트레스를 높이고, 이로 인해 단백질 분해 활성화가 더욱 촉진되어 단백질 항상성 손상이 가속화된다(Ulla 등, 2021; Zeng 등, 2024).
이러한 병태생리적 기전을 표적으로 한 근감소증을 예방하는 효능을 가진 다양한 천연물에 대한 연구가 활발히 진행되고 있다. Dex로 근위축을 유도시킨 C2C12 세포 및 C57BL/6 마우스 모델에서 구절초(Chrysanthemum zawadskii Herbich) 추출물은 근육 손실과 기능 저하를 유의하게 완화하는 것으로 보고되었다(Lee 등, 2021). 이러한 구절초의 효과는 GR의 핵 내 전위 억제를 통해 Atrogin-1 및 MuRF1 유전자 발현을 감소시키고, Akt/mTOR 신호 활성화를 통한 단백질 항상성 조절과 더불어 단백질 분해 및 미토콘드리아 기능 이상을 억제하는 데 기인한다. 한편, 길초근(Valeriana fauriei) 추출물 또한 동일 모델에서 근위축을 억제하였으며, ROS 생성 억제를 통해 산화 스트레스를 완화함으로써 Atrogin-1, MuRF1, myostatin 등의 유전자 발현을 감소시키고, GR 및 FoxO3α의 핵 내 전위 억제를 통해 근위축 완화 효과를 나타냈다(Kim 등, 2022). 또한, 물쑥(Peucedanum japonicum Thunb) 추출물 역시 in vitro와 in vivo 모델에서 Dex에 의한 근위축을 예방하였다(Kim 등, 2024). 물쑥 추출물은 GR과 FoxO3α의 핵 내 전위를 저해하고 MuRF1과 Atrogin-1의 전사를 억제함으로써 단백질 분해 기전을 억제하였다. 반면 근육 분화 관련 지표인 myogenin (Myog), myogenic differentiation 1 (Myod1)의 유전자 발현을 증가시키고 Akt/mTOR 신호 및 그 하위 신호인 ribosomal S6 kinase (S6K)와 eukaryotic translation initiation factor 4E binding protein 1 (4EBP1)의 활성화를 통해 단백질 합성을 높여 근섬유 형성을 촉진하였다.
천연물의 유효 성분이 GR 신호기전을 통해 근감소증 완화 효과를 보인다는 연구결과도 다양하게 보고되었다. 인삼과 홍삼의 주요 활성 사포닌 성분인 진세노사이드(ginsenoside)는 Dex로 유도한 근위축 C2C12 세포 및 C57BL/6N 마우스 모델에서 ROS 감소 및 미토콘드리아 기능 개선을 비롯한 산화 스트레스 억제, 그리고 염증성 사이토카인과 섬유화 관련 신호인 Transforming Growth Factor-β (TGF-β) 경로 억제 등의 염증 반응 감소를 통해 근육 위축을 완화하였다(Kim 등, 2025). 또한 in silico molecular docking 분석 결과, 진세노사이드가 GR에 경쟁적으로 결합하여 GR 신호 전달을 저해하는 것으로 확인되었다. 한편, 천연 펩타이드인 카르노신(carnosine)도 Dex로 유도된 근위축 C57BL/6J 마우스 모델에서 ROS를 감소시켜 산화 스트레스에 민감한 ubiquitin ligase인 casitas B-lineage lymphoma proto-oncogene-b (Cbl-b)와 GR 관련 단백질 분해 유전자의 발현을 억제함으로써 근육량과 근기능을 보존하는 효과를 보였다(Rahman 등, 2025). 식물 유래 플라보노이드인 모린(morin) 투여 역시 동일 모델에서 Klf15, FoxO3a,MuRF1 등 GR 하위 신호를 조절하고, Cbl-b, nuclear factor erythroid 2-related factor 2, SOD1과 같은 산화 스트레스 관련 지표를 조절함으로써 근육 단백질 손상과 기능 저하를 억제하였다(Ulla 등, 2022). 이처럼 다양한 천연물이나 유효 성분이 GR 매개 전사 작용을 조절하여 염증성 사이토카인 발현을 억제하거나 산화 스트레스를 감소시킴으로써 근육 세포의 손상 및 기능 저하를 억제하여 근감소증 완화에 긍정적인 영향을 미칠 수 있다.
꾸지뽕나무의 학명은 Cudrania tricuspidata이며 동아시아(한국, 중국, 일본 등)에 분포하는 낙엽관목류로, 전통적으로 잎과 뿌리, 열매 등이 식품 및 민간약재로 널리 활용되어 왔다(Xin 등, 2017). 꾸지뽕나무잎에는 kaempferol, quercimeritrin, cycloartocarpesin 등 다양한 폴리페놀(polyphenol) 및 플라보노이드(flavonoid) 계열 화합물이 풍부하게 함유되어 있으며, 이들 성분은 항산화, 항염증 및 항당뇨 활성 등 다양한 생리활성을 갖는 것으로 보고되었다(Lee 등, 2011; Lee 등, 2024; Xin 등, 2017). 특히, Jo 등(2021)은 꾸지뽕나무잎에서 분리한 xanthone 및 플라보노이드 유도체가 우수한 항산화 활성과 항-티로시나제(anti-tyrosinase) 활성을 나타냄을 보고하여, 꾸지뽕나무잎이 산화적 손상 억제에 기여할 수 있음을 제시하였다(Jo 등, 2021a). 또한, kaempferol은 ROS를 효과적으로 제거하고, 항산화 효소인 SOD, CAT, GPx의 발현을 증가시켜 세포 내 산화적 손상을 완화하는 것으로 알려져 있다(Speisky 등, 2023). 한편, 꾸지뽕나무잎 추출물은 lipopolysaccharide (LPS)로 유도된 염증세포에서 nitric oxide (NO) 및 염증성 사이토카인인 tumor necrosis factor-alpha (TNF-α)와 interleukin-6 (IL-6) 등의 생성을 억제하였다(Ko 등, 2023). 이는 nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) 및 mitogen-activated protein kinase (MAPK) 신호전달 경로를 조절함으로써 항염증 효과를 나타낸다고 보고되었다. 더불어, 꾸지뽕나무잎에서 분리된 프레닐화 이소플라보노이드는 α-glucosidase 억제 및 단백질 당화 저해 활성을 통해 혈당 조절과 대사 기능 개선 효과를 나타내는 것으로 보고되었다(Jo 등, 2021b). 이러한 연구 결과들은 꾸지뽕나무잎이 산화 스트레스 및 염증으로 유도되는 질환을 예방하거나 치료하는데 기여할 수 있음을 시사한다. 따라서 본 연구에서는 항산화, 항염증 등의 활성을 지닌 꾸지뽕나무잎의 근위축 예방 효과와 관련 분자 기전을 Dex로 근위축을 유도한 C2C12 세포 모델에서 규명하고자 하였다.
2. 재료 및 방법
Dulbecco’s modified Eagle’s medium (DMEM), penicillin-streptomycin, fetal bovine serum (FBS), horse serum (HS), phosphate-buffered saline (PBS), Dulbecco’s phosphate-buffered saline (DPBS) 및 0.25% trypsin-ethylenediaminetetraacetic acid (EDTA)는 WELGENE (Daegu, Korea)에서 구입하였다. 1차 항체인 Myosin heavy chain (MyHC), HSC70, MuRF1은 Santa Cruz Biotechnology Inc. (Dallas, TX, USA)에서 구입하였고, p-mTOR과 p-Akt는 Cell Signaling Technology (Beverly, MA, USA)에서 구입하였다. Horseradish peroxidase (HRP)-conjugated 2차 항체는 Jackson Immuno Research Laboratories (West Grove, PA, USA)에서 구입하였다. Dexamethasone (Dex), dimethyl sulfoxide (DMSO), chlorogenic acid (≥95.0%), isoquercetin (≥90%), gallic acid, catechin, 2,2-diphenyl-1-picrylhydrazyl (DPPH), 6-hydroxy-2,5,7,8-tetramethylchromane-2-carboxylic acid (Trolox), aluminum chloride hexahydrate 및 Folin-Ciocalteu 용액은 Sigma-Aldrich (St. Louis, MO, USA)에서 구입하였다. Kaempferol 3-O-glucoside (100%)는 한국천연물과학기술연구소(Anseong, Korea)에서 분양받았다. Acetonitrile (ACN)과 methanol (MeOH)은 Fisher Scientific Inc. (Cleveland, OH, USA)에서 구입하였다. 아스코르빈산(ascorbic acid)과 2,2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS)는 각각 TCI (Tokyo, Japan), Kanto Chemical (Tokyo, Japan)에서 구입하였다. Potassium persulfate은 Junsei (Tokyo, Japan)에서, sodium carbonate는 Samchun Pure Chemical Co., Ltd. (Seoul, Korea)에서, sodium nitrite solution 및 sodium hydroxide는 Duksan Pure Chemicals Co., Ltd. (Jincheon, Korea)에서 구입하여 사용하였다.
본 실험에 사용된 꾸지뽕나무잎은 경상북도 영천시에 위치한 농업회사법인 ㈜두손애약초에서 구매하였다. 구매한 시료는 수세한 후 진공동결건조기(Model FD8508, ilShinBioBase Co., Ltd., Dongducheon, Korea)를 사용하여 -80°C, 5 mTorr의 감압 조건에서 동결건조한 후 분말화하여, 70% 에탄올에 10% w/v 농도로 혼합하고 실온에서 24시간 동안 추출하였다. 추출물은 면포와 여과지로 한 번씩 여과한 후 rotary vacuum evaporator (EYELA, Tokyo, Japan)를 이용해 농축한 뒤 다시 동결 건조하였다. 분쇄하여 -20°C에 보관하였고, 사용 직전 DMSO에 녹여 1,000배 농도의 stock 용액을 제조하였다.
C. tricuspidata leaf extract (CTE)의 DPPH radical 소거 활성은 Brand-Williams의 방법을 변형하여 측정하였다(Brand-Williams, 1995). 각 well 당 CTE은 50 μL씩, 0.02% DPPH 메탄올 용액은 100 μL씩 넣고 혼합하여 암실에서 30분간 반응시켰다. SpectraMax® ID3 (Molecular Devices, San Jose, CA, USA)을 이용하여 510 nm에서 흡광도를 측정하였다. 기존에 알려진 항산화제인 ascorbic acid를 표준물질로 사용하여 얻은 표준곡선은 대조군을 사용하여 CTE의 DPPH radical 소거 활성을 구하였으며, 단위는 milligram ascorbic acid equivalent (mg AAE)/mL를 사용하였다.
CTE의 ABTS radical 소거 활성은 Biglari의 방법을 변형하여 측정하였다(Biglari 등, 2008). ABTS 혼합 용액은 7.4 mM ABTS와 2.6 mM potassium persulfate를 1:1로 혼합한 후 암실에서 16시간 동안 반응시킨 후 SpectraMax® ID3를 사용해 734 nm에서의 흡광도 값이 0.70±0.02가 되도록 증류수로 희석하여 ABTS 용액을 제조하였다. 이후, CTE 10 μL와 ABTS 용액 190 μL를 혼합하여 암실 상태의 상온에서 6분간 반응시킨 후 732 nm에서 흡광도를 측정하였다. Trolox를 표준물질로 사용하여 얻은 표준곡선을 대조군으로 사용하여 CTE의 ABTS radical 소거 활성을 구하였으며, 단위는 μM Trolox equivalent (μM TE)/mL를 사용하였다.
CTE의 총폴리페놀 함량은 Folin-Ciocalteu 방법을 변형하여 분석하였다(Singleton 등, 1999). 각 농도의 CTE 15 μL에 30 μL의 10% Folin-Ciocalteu 용액을 혼합한 후 암실에서 5분간 반응시켰다. 그 후 120 μL의 700 mM sodium carbonate를 첨가하여 상온에서 1시간 동안 암실에서 반응시킨 후 765 nm에서 흡광도를 측정하였다. 표준물질로 95% 메탄올에 녹인 gallic acid를 사용하여 얻은 표준곡선을 대조군으로 사용하여 CTE의 총폴리페놀 함량을 구하였으며, 단위는 milligram gallic acid equivalent (mg GAE)/mL를 사용하였다.
CTE의 총플라보노이드 함량은 aluminum chloride colorimetric method를 변형하여 분석하였다(Zhishen 등, 1999). 각 농도의 CTE 25 μL에 증류수 125 μL와 5% sodium nitrite solution 10 μL를 혼합하여 6분간 상온에서 반응시킨 후, 10% aluminum chloride hexahydrate 15 μL를 추가하여 상온에서 5분간 반응시켰다. 그 후 1 M sodium hydroxide 50 μL와 증류수 125 μL를 혼합하여 510 nm에서 흡광도를 측정하였다. 표준물질로 증류수에 희석한 catechin을 사용하여 얻은 표준곡선으로부터 CTE의 총플라보노이드 함량을 구하였으며, 단위는 milligram catechin equivalent (mg CE)/mL를 사용하였다.
본 실험에 사용된 C2C12 myotube (CRL-1772, American Type Culture Collection, VA, USA)는 마우스 유래 골격근 세포주로 10% FBS-DMEM을 사용하여 37°C, 5% CO2 조건으로 90 mm cell culture dish (SPL Life-Sciences, Pocheon, Korea)에서 배양하였다. 분화 유도를 위해 세포를 6-well 또는 24-well plate에 계대 배양한 후, 세포 밀도가 약 90%에 도달하면 2% HS-DMEM으로 배지를 교체하여 6일간 분화를 유도하였다. 분화 6일차에 100 μM Dex와 다양한 농도(100, 200, 300 μg/mL)의 CTE를 24시간 병용 처리하였다.
CTE의 세포 독성을 확인하기 위하여 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) 분석법을 사용하여 세포 생존율을 측정하였다. C2C12 세포를 96-well cell culture plate (SPL Life Science)에 0.1×105 cells/well의 농도로 분주한 후, 37°C, 5% CO2 조건에서 24시간 배양하였다. 세포 밀도가 약 80-90%에 도달하면 CTE (100, 200, 300 μg/mL)를 단독으로 처리하거나 100 μM Dex와 병용 처리하였다. 24시간 처리 후 상등액을 제거하고, 10% FBS-DMEM 배지에 5 mg/mL의 MTT 용액을 10%(v/v)가 되도록 희석하여 각 well에 첨가한 뒤, 37°C, 5% CO2 조건에서 3시간 동안 반응시켰다. 반응 종료 후 상등액을 제거하고, 생성된 남색 formazan 결정을 DMSO로 완전히 용해시킨 다음 570 nm에서 흡광도를 측정하였다. 측정된 흡광도는 % of control로 표현하였다.
24시간 시료 처리한 C2C12 세포는 PBS로 2회 세척한 후, 100% MeOH 고정 및 자연 건조하였다. 이후 Jenner 염색 용액(Electron Microscopy Sciences, Hatfield, PA, USA)과 PBS를 1:3 비율로 희석하여 5분간 반응시킨 후, Giemsa stain (Sigma-Aldrich)과 증류수를 1:20 비율로 희석하여 10분간 반응시켜 염색하였다. 염색된 세포는 형광 현미경(Eclipse TS2-LS, Nikon, Tokyo, Japan)을 이용하여 촬영하였으며, NIS-Element 분석 소프트웨어(Version 5.30.03, Nikon, Japan)으로 myotube 길이와 직경을 측정하여 정량화하였다.
시료 처리한 C2C12 세포를 DPBS로 세척한 후 3% paraformaldehyde (Samchun Pure Chemical Co., Ltd.)로 15분간 고정시켰다. 고정된 세포에 0.1% saponin (Sigma-Aldrich)를 분주하여 10분간 투과시킨 후 1% bovine serum albumin (BSA; Bovogen, Australia)로 상온에서 1시간 블로킹(blocking)하였다. 이후 MyHC 1차 항체(1:500)를 4°C에서 하룻밤(overnight) 반응시킨 뒤 Alexa Fluor 488-conjugated anti-mouse IgG (1:1,000; Cell Signaling Technology)를 상온에서 1시간 반응시켰다. 마지막으로 1 μg/mL 4′,6-diamidino-2-phenylindole (DAPI; Roche, CA, USA)를 5분간 처리하였다. 염색된 세포는 형광 현미경을 이용하여 MyHC 양성 근관을 촬영하였다.
총RNA는 RNAiso Plus (Takara, Kyoto, Japan)를 이용하여 추출하였으며, chloroform (99%, Extra Pure, Duksan Pure Chemicals) 처리 후 원심 분리하여 상층액을 회수하였다. 상층액에 iso-propyl alcohol (Duksan Pure Chemicals)을 첨가하여 다시 원심 분리한 뒤, 생성된 pellet은 75% 에탄올로 세척하여 정제하였다. RNA 농도 및 순도는 NanoDrop Lite spectrophotometer (UL 61010-1, Thermo Fisher Scientific, Waltham, MA, USA)를 이용해 260/280 nm에서 측정하였으며, A260/A280 비율이 1.8-2.0인 RNA만 cDNA 합성에 사용하였다. cDNA 합성은 AMPIGENE® cDNA Synthesis Kit (Enzo Life Sciences, Farmingdale, NY, USA)를 제조사 지침에 따라 합성하였다. qRT-PCR은 AMPIGENE® qPCR Green Mix Hi-ROX (Enzo Life Sciences, Farmingdale, NY, USA)를 사용하여 20 μL 반응 용량으로 수행하였다. 증폭 조건은 95°C에서 10초(변성), 60°C에서 15초(결합), 72°C에서 15초(신장) 과정을 40회 반복하였다. 18S rRNA를 기준 유전자로 사용하였으며, 상대 유전자 발현량은 2-ΔΔCt 방법으로 계산하였다. 실험에 사용된 프라이머(primer)의 염기서열은 Redd1 (forward, 5′-CCT AGC CTC TGG GAT CGT TTC-3′; reverse, 5′-ATC AGC GGC CGG AGT TC-3′), Klf15 (forward, 5′-TGG TAC CAT CCT CCA ACT TGA A-3′; reverse, 5′-CAA TAG GTT TGG CGG CAA TG-3′)이며, BIONEER (Daejeon, Korea)에서 합성하였다.
C2C12 세포는 DPBS로 세척 후 회수하여 lysis buffer (100 mM Tris-HCl, 100 mM NaCl, 0.5% Triton X-100, 1 mM sodium orthovanadate, 10 mM sodium fluoride, protease inhibitor cocktail)로 용해하였다. 원심 분리하여 얻은 상층액의 단백질 농도는 Bradford solution (Bio-Rad Laboratories, Hercules, CA, USA)을 이용하여 Bradford assay로 측정하였다. 동일한 양의 단백질을 sodium dodecyl sulfate-polyacrylamide gel으로 분리하여 polyvinylidene difluoride membrane로 전이한 후, 5% BSA로 블로킹하고 1차 항체(1:1,000)를 4°C에서 overnight 반응시켰다. 그 후 1x Tris-buffered saline-Tween 20 (10 mM Tris-HCl, 150 mM NaCl, 0.05% Tween-20)으로 세척하고 HPR-conjugated anti-mouse 또는 anti-rabbit 2차 항체(1:5,000)를 실온에서 2시간 반응시켰다. 단백질 밴드는 enhanced chemiluminescence (ECL) detection reagent (Thermo Fisher Scientific Inc.)으로 검출하고 ChemiDoc imaging system (Model 300; Azure Biosystems, Dublin, CA, USA)으로 시각화하였다. 밴드 강도는 ImageJ software (NIH, Bethesda, MD, USA)로 정량분석하였다.
CTE는 Diode Array Detector가 장착된 HPLC 시스템(1200 series, Agilent Technologies, California, USA)을 사용하여 수행되었다. 시료는 27°C에서 Eclipse XDB-C18 컬럼(250×4.6 mm, 5 μm)을 이용하여 분리하였다. 이동상은 0.1% phosphoric acid(용매 A)와 100% ACN(용매 B)로 구성되었다. 기울기 용출(gradient elution)은 다음과 같은 시간 프로그램에 따라 진행되었다: 0-3분, 용매 B 5-12%; 3-8분, 용매 B 12-14%; 8-25분, 용매 B 14-20%; 25-35분, 용매 B 20-35%; 35-45분, 용매 B 35-60%; 45-60분, 용매 B 60-100%; 60-65분, 용매 B 100-5%. 모든 시료는 0.45 μm 멤브레인 필터로 여과 후, 10 μL를 주입하고 유속은 0.8 mL/min로 설정하였다. Chlorogenic acid의 흡광도는 320 nm에서, isoquercetin과 kaempferol-3-glucoside의 흡광도는 360 nm에서 측정하였다. 각 화합물의 정량은 표준품으로 작성한 검량곡선을 기반으로 하였으며, 결과는 μg/g of extract 단위로 표기하였다.
GR의 결정구조(PDB ID:1M2Z)는 PDB 데이터베이스 (https://www.rcsb.org/)에서 수집하였다. Ligand로 사용된 CTE 내 3종의 폴리페놀(chlorogenic acid, isoquercetin, kaempferol-3-glucoside)의 3차원 화학 구조는 PubChem 데이터베이스(https://pubchem.ncbi.nlm.nih.gov/)에서 내려받았다. GR 및 ligand로 사용된 폴리페놀은 OpenBabel 소프트웨어(v2.4.1)를 사용하여 PDB 형식에서 도킹에 적합한 PDBQT 파일로 변환하였다. 분자 도킹은 AutoDock Vina (v1.5.7)을 사용해 수행하였으며, 도킹 스코어로 결합 친화도를 평가하였다. CTE 내 폴리페놀 3종 중 결합 친화도가 가장 높은 2종의 도킹 포즈를 선정하여 BIOVIA Discovery Studio Client (2024)를 통해 3차원 구조 및 2차원 상호작용 다이어그램으로 시각화하였다.
3. 결과 및 고찰
CTE의 농도별 항산화능(DPPH 라디칼 소거능, ABTS 라디칼 소거능), 총폴리페놀(total phenolic contents, TPC)과 총플라보노이드 함량(total flavonoid contents, TFC)을 측정한 결과를 Table 1에 제시하였다. CTE의 DPPH 및 ABTS 라디칼 소거능은 농도 의존적으로 증가하였으며, 가장 높은 농도인 5 mg/mL에서 각각 0.050±0.001 mg AA/mL 및 994.94±17.20 μM TE/mL의 활성을 나타냈다. TPC와 TFC의 결과 또한 이와 유사한 농도 의존적 경향을 확인하였으며, 각각 최고 농도인 5 mg/mL에서 0.194±0.001 mg GAE/mL 및 0.184±0.000 mg CE/mL 값을 보였다.
항산화 활성은 시료의 TPC 및 TFC와 밀접한 상관관계를 갖는 것으로 알려져 있다. Dai와 Mumper (2010)는 다양한 식물 추출물과 식품 성분을 대상으로 한 연구에서 TPC와 TFC가 높을수록 DPPH 및 ABTS 라디칼 소거능 등의 항산화 지표가 유의하게 증가한다는 문헌적 ․ 실험적 근거를 리뷰하였으며, 특히 페놀 및 플라보노이드 화합물이 자유라디칼 소거능과 금속 이온 킬레이션을 통해 항산화 활성에 기여하는 핵심 인자임을 보고하였다(Dai과 Mumper, 2010). 또한, Muflihah 등(2021)은 12종 인도네시아 허브 추출물의 TPC와 TFC 값이 DPPH 라디칼 소거능 및 FRAP 활성 간에 0.92-0.97의 매우 높은 상관계수를 나타냄으로써, 이들 성분이 항산화 활성을 예측하는 주요 지표임을 통계적으로 입증하였다(Muflihah 등, 2021). 본 연구에서도 이러한 선행 연구와 일치하게, CTE 농도가 높을수록 TPC와 TFC가 함께 증가하는 경향을 확인하였다. 따라서, CTE의 항산화 능력은 TPC와 TFC에 기인하는 것으로 사료된다.
CTE의 유효성분을 확인하고자 HPLC 분석을 수행하였다. 분석 결과, CTE에는 chlorogenic acid, isoquercetin, 그리고 kaempferol-3-glucoside가 포함되어 있는 것으로 확인되었다(Table 2). 함량은 chlorogenic acid가 326.25 μg/g으로 가장 많았고 그 다음 순으로 isoquercetin은 41.36 μg/g, kaempferol-3-glucoside는 30.19 μg/g순으로 확인되었다.
이러한 성분조성은 선행연구에서 보고된 CTE의 주요 생리활성 성분과 유사한 경향을 보였다. Seo 등(2022)은 CTE에서 isoquercetin 및 kaempferol-3-glucoside를 주요 성분으로 확인하였으며(Seo 등, 2022), Song 등(2017)은 chlorogenic acid와 kaempferol을 HPLC 분석을 통해 검출한 바 있다(Song 등, 2017). 반면, Choi 등(2023)은 protocatechuic acid, catechin, cryptochlorogenic acid, caffeic acid, epicatechin 등 본 연구 시료에서는 검출되지 않은 화합물을 보고하였다(Choi 등, 2023). 이는 본 연구와 선행연구에서 사용한 추출 조건 및 HPLC 분석 조건의 차이 때문으로 판단된다. 천연물은 채취 지역 및 시기에 따라 차이가 존재할 수 있어 시료 표준화를 위해서 향후 다양한 채취 지역 및 시기의 꾸지뽕나무잎에 대한 성분 및 함량 분석 연구가 추가적으로 필요할 것으로 사료된다.
CTE의 세포 독성을 평가하기 위해 MTT 분석을 수행하였다. C2C12 세포에 Dex를 처리하지 않은 상태에서 CTE를 다양한 농도(100, 200, 300 μg/mL)로 단독 처리한 결과, 가장 높은 농도(300 μg/mL)에서도 세포 생존율은 대조군과 유사하게 유지되어 CTE가 세포 독성을 유발하지 않음을 확인하였다(Fig. 1A). 반면, C2C12 세포에 100 μM의 Dex를 단독 처리하였을 때 세포 생존율은 대조군 대비 80% 이하로 감소하였으며, 동일한 농도의 Dex와 CTE를 병용 처리한 군에서도 세포 생존율은 Dex 처리군과 유사한 수준으로 유지되었다(Fig. 1B). 따라서 Dex와 CTE를 병용 처리한 군에서 관찰된 세포 생존율 감소는 Dex 처리에 의한 영향이며 CTE의 추가적인 세포독성은 없는 것으로 판단된다.
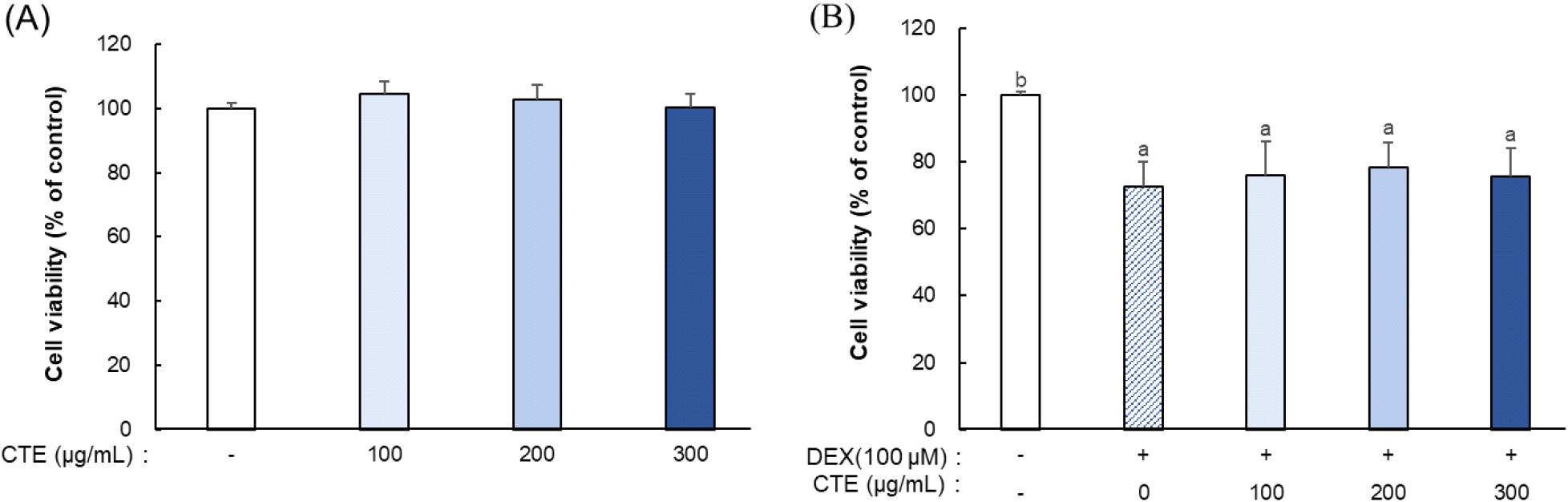
Dex는 C2C12 세포의 생존율을 감소시키는 주요 약물로 알려져 있다. Dex를 농도별(125, 250, 500, 1,000 μM)로 C2C12 세포에 4시간 동안 처리한 결과, 가장 낮은 처리 농도인 125 μM에서부터 세포 생존율이 대조군 대비 80% 이하로 유의하게 감소되었음을 확인하였다(Chen 등, 2020). 또한, 세포 생존율은 농도 의존적으로 감소하여, 최고 농도인 1,000 μM Dex 처리군에서는 50% 이하로 낮아졌다. Dex 처리는 ROS 생성을 유도하고 BCL2-associated X protein (Bax)/B-cell lymphoma 2 (Bcl-2) 비율을 증가시킨다. 이는 미토콘드리아의 막 투과성을 증가시켜 cytochrome c (Cyt c) 방출을 초래하고, 방출된 Cyt c는 apoptotic peptidase activating factor 1 (Apaf-1)과 결합하여 apoptosome을 형성한다. 최종적으로 이 apoptosome은 caspase-9 및 caspase-3을 순차적으로 활성화시키며 내재성 apoptosis 경로를 통해 세포사멸을 유도하는 것으로 보고되었다. 본 연구에서 사용한 Dex 농도(100 μM)에서도 이와 유사한 세포 생존율 감소가 확인되어, Chen 등(2020)에서 보고한 Dex 처리에 기인한 세포사멸과 유사한 양상으로 판단된다. 따라서 이후 실험에서는 100, 200, 300 μg/mL의 CTE 농도를 적용하여 Dex 유도 근위축 모델에서 CTE의 보호 효과를 검증하였다.
Dex로 유도된 근관 위축에 대한 CTE의 억제 효과를 평가하기 위해, 분화가 완료된 C2C12 근관세포의 형태학적 변화를 관찰하였다. 세포는 Dex 100 μM 단독 또는 Dex와 CTE를 농도별(100, 200, 300 μg/mL)로 24시간 동안 병용 처리한 후, Jenner-Giemsa 염색을 실시하여 근관의 직경과 길이를 측정하였다(Fig. 2A). 그 결과, Dex 처리군에서는 근관의 직경과 길이가 대조군(control)에 비해 각각 약 55.45%, 35.15% 감소하여 근위축이 뚜렷하게 나타났다. 반면, CTE를 병용 처리한 군에서는 Dex에 의한 근관 위축이 농도 의존적으로 완화되었다. 특히 300 μg/mL CTE 처리군에서는 Dex 처리군 대비 근관의 직경이 136.61%, 길이가 36%로 각각 유의하게 증가하였다. 이를 재확인하기 위하여 MyHC 양성 근관의 발현 수준을 형광염색법으로 평가했다(Fig. 2B). 대조군에 비해 Dex 처리군에서 근관 직경과 MyHC 형광 발현이 모두 감소하는 것이 확인되었다. 또한 Dex 처리군과 비교하여 CTE 처리군은 Dex 처리로 인해 감소했던 MyHC의 양성 근관의 발현 수준을 증가시켰을 뿐만 아니라, 근관 직경도 증가시키는 효과를 보였다.
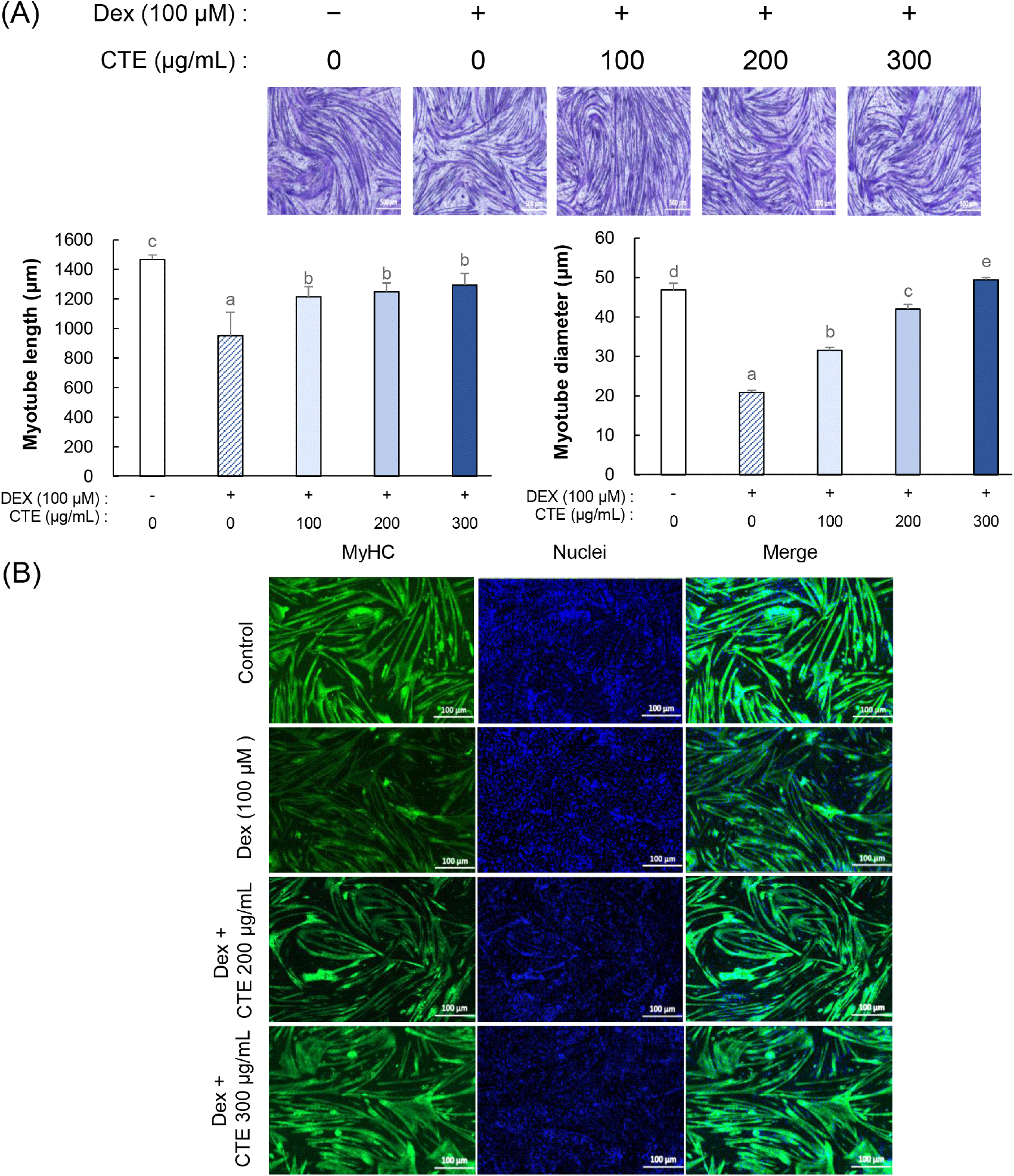
선행 연구에서 천연물 유래 유효성분들이 근위축을 예방하며 MyHC 단백질 발현을 촉진함을 확인하였다. MyHC는 근육 수축에 관여하며, 새로운 근섬유 형성뿐만 아니라 기존 근섬유의 유지 및 성장에 필수적인 구조 단백질이다(Schiaffino, 2015). 따라서 MyHC 발현의 감소는 근위축을 나타내는 대표적인 지표로 알려져 있다. 예를 들어, Xing 등(2021)은 Wistar rats을 2주간 비활동 상태로 유지하여 근위축이 유도된 동물 모델에 chlorogenic acid를 급여한 결과, 산화 스트레스를 저하시켜 근섬유 크기 감소를 억제했다고 보고하였다(Xing 등, 2021). 또한, Mandal 등(2025)은 quercetin과 kaempferol의 단독 및 혼합물 처리가 lipopolysaccharide (LPS)로 유도된 C2C12 근위축 모델에서 MyHC의 발현 저하 및 근관의 직경 감소를 완화한다고 보고하였다(Mandal 등, 2025). ICR 마우스 모델에서 2주간의 isoquercitrin 급여는 탈신경으로 유도된 가자미근의 위축과 MyHC 발현 감소를 유의하게 완화하는 것으로 보고되었다(Shen 등, 2020). 한편, p-coumaric acid의 투여는 고지방 및 고과당 식이로 근위축을 유도한 C57BL/6J 마우스 모델에서 감소된 MyHC 발현을 유의하게 증가시켜 근위축 완화에 기여한 것으로 보고되었다(Truong 등, 2024).
Fig. 2에서 관찰된 근관의 형태학적 변화와 MyHC의 발현에 대한 CTE의 예방 효과를 검증하기 위해, 단백질 항상성 관련 지표의 발현을 확인하였다. MyHC 단백질 발현은 Dex 처리군에서 대조군 대비 약 0.58배 수준으로 유의적으로 감소하였다(Fig. 3A). 반면 CTE 처리군은 100 μg/mL에서는 Dex 처리군과 유의한 차이를 보이지 않았으나, 200 μg/mL에서 약 1.67배 증가하여 대조군과 유사한 발현 수준을 회복하였다. 최고 농도인 300 μg/mL에서는 Dex 처리군 대비 약 2.1배 증가하여 대조군 수준을 상회하였다. 다음으로 단백질 합성과 관련된 주요 지표인 p-mTOR와 p-Akt의 발현을 확인하였다. p-mTOR의 발현은 Dex 처리군에서 대조군 대비 약 0.24배로 유의하게 감소하였으나, CTE 처리군에서는 Dex 처리군 대비 농도 의존적으로 증가하여 100 μg/mL에서 약 1.95배, 200 μg/mL에서 약 2.4배, 300 μg/mL에서 약 3.59배로 상향 조절되었다. 한편, p-Akt의 단백질 발현은 Dex 처리군에서 대조군 대비 약 0.46배 감소하였지만, CTE 처리군에서는 Dex 처리군 대비 100 μg/mL에서 약 1.72배, 200 μg/mL에서 약 2.25배 증가하였다. 최고 농도인 300 μg/mL에서는 약 2.2배 증가하였지만, 200 μg/mL 처리군과 통계적으로 유의한 차이는 관찰되지 않았다. 반면, 단백질 분해 기전의 주요 지표인 MuRF1의 발현은 Dex 처리군에서 대조군 대비 약 2.83배 증가한 것으로 확인되었다(Fig. 3B). CTE 처리 시 Dex 처리군 대비 농도 의존적으로 감소하여, 100 μg/mL에서 약 0.79배, 200 μg/mL에서 약 0.69배, 300 μg/mL에서 약 0.39배 수준으로 유의하게 억제되었다.
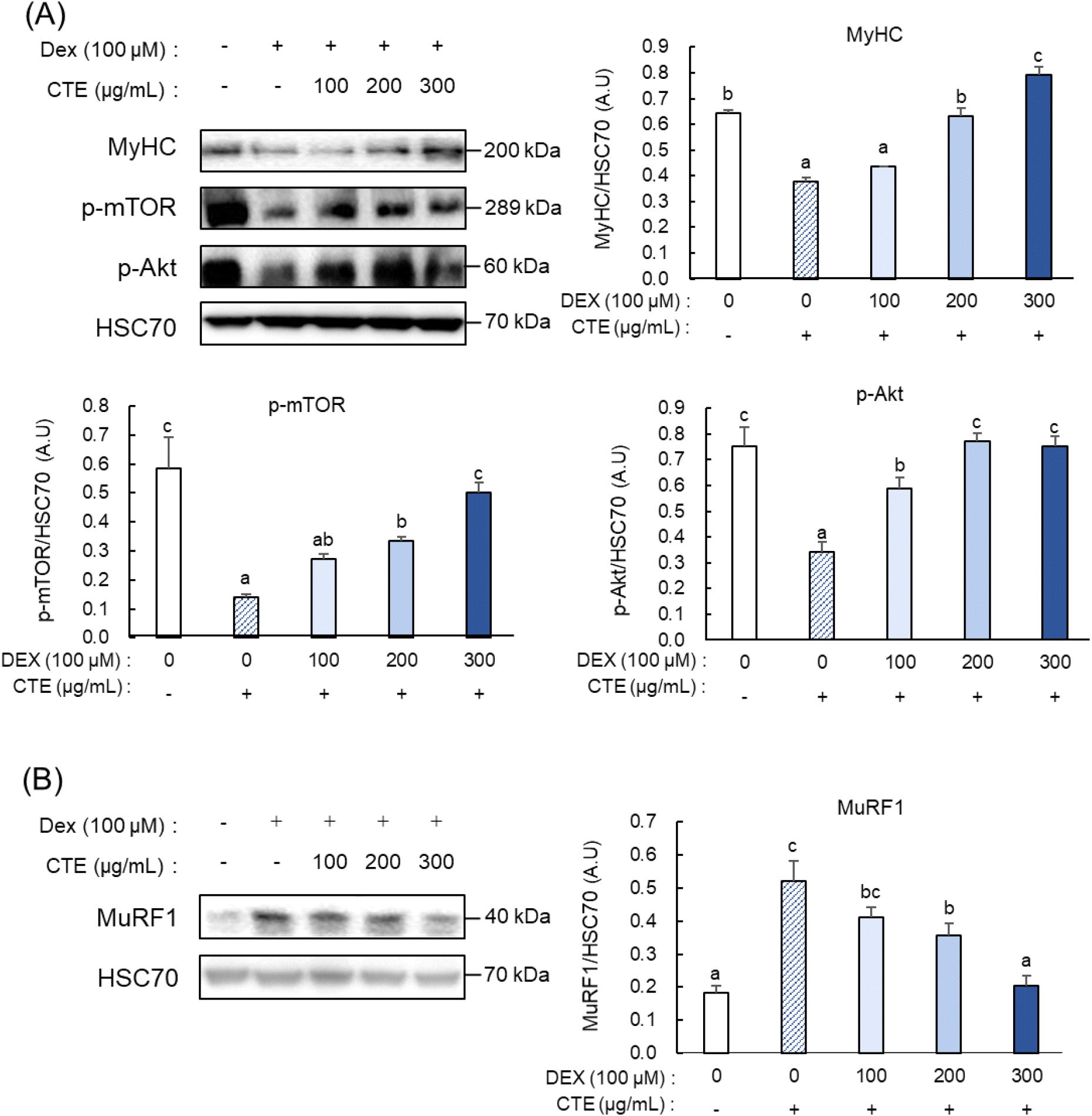
단백질 항상성의 불균형은 근감소증의 주요 원인으로 잘 알려져 있다(Kim 등, 2023b). 이를 조절하는 주요 경로 중 하나인 Akt/mTOR 신호전달 경로의 활성화는 근육 단백질 합성을 촉진하고 골격근의 성장 및 재생을 지원하는 데 필수적인 역할을 한다(Morita 등, 2015). Zhang 등(2024)은 C2C12 근관세포에 15종의 필수 및 분지쇄 아미노산(branched chain amino acid)의 일정 비율 혼합물을 처리했을 때 Akt가 활성화되어 mTOR 신호전달 경로를 자극하고, 이에 따라 하위 신호인 S6K의 인산화가 증가하여 단백질 합성이 촉진됨을 확인하였다(Zhang 등, 2024). 반면, 유비퀴틴-프로테아좀 시스템(ubiquitin-proteasome system) 내에서 MuRF1의 과도한 발현은 단백질 분해를 촉진하여 근위축을 유발하는 주요 기전으로 보고되고 있다(Bodine과 Baehr, 2014). Park 등(2017)에 따르면, C2C12 근관 세포에 Dex와 hydrogen peroxide를 처리했을 때 MuRF1과 Atrogin-1의 발현이 대조군에 비해 유의하게 증가하였으며, 이는 유비퀴틴-프로테아좀 시스템 경로의 활성화로 근위축이 유도되었음을 시사한다(Park 등, 2017). 이처럼 단백질 항상성의 불균형은 근위축의 주요 원인으로 작용하며, 이를 타겟으로 근위축을 개선한 in vivo 연구도 보고된 바 있다. He 등(2024)은 5/6 신절제술로 만성 신장질환성 근위축이 유도된 Sprague-Dawley rat에 caffeic acid를 10주간 경구 투여한 결과, 단백질 합성 지표인 p-Akt 및 p-mTOR의 발현이 회복되고, 단백질 분해 지표인 Atrogin-1과 MuRF1의 발현이 유의하게 억제됨을 확인하였다(He 등, 2024). 이는 Akt/mTOR 경로 활성화와 유비퀴틴-프로테아좀 시스템 억제가 동시에 이루어짐으로써 근위축이 개선됨을 시사한다.
따라서 본 연구의 결과는 CTE가 단백질 합성을 촉진하고 단백질 분해를 억제함으로써, Dex로 유도된 근관세포의 단백질 항상성 불균형을 회복하고 근위축을 완화할 수 있음을 보여준다.
CTE에서 확인된 유효 성분이 근위축 완화에 어떻게 기여하는지 그 작용 기전을 탐색하기 위해 molecular docking 분석을 수행하였다. HPLC 분석을 통해 확인된 CTE 내 주요 폴리페놀 3종과 GR 간의 결합 친화도를 평가한 결과, chlorogenic acid가 -7.17 kcal/mol로 가장 강한 결합 친화도를 보였고 kaempferol-3-glucoside도 -7.04 kcal/mol의 비교적 높은 결합 친화도를 나타냈다(Fig. 4A). 또한, isoquercetin도 -6.45 kcal/mol로 안정적인 결합 친화도를 확인하였다. 결합 모드 분석에서 chlorogenic acid는 GR 결합 포켓 내에서 Met604, Phe623, Arg611, Gln570, Asn564와의 수소결합 및 Leu566, Leu608과의 π-alkyl 상호작용, 그리고 van der Waals 접촉 등을 형성하여 안정적으로 결합하였다. Kaempferol-3-glucoside 역시 Arg614, Gln615, Glu537, Arg611, Trp610 및 Pro541과의 수소결합, Tyr660과의 π-π T-shaped stacking, Val543과의 π-alkyl 상호작용, 그리고 여러 잔기와의 van der Waals 접촉 등을 통해 포켓 내부에서 안정적인 결합 양상을 보였다. 다만, chlorogenic acid와 kaempferol-3-glucoside 모두 unfavorable donor-donor 상호작용이 관찰되었다. 이러한 in silico 분석 결과를 검증하기 위하여 GR 하위 전사인자의 발현을 확인하였다(Fig. 4B). CTE는 근육 단백질 분해에 기여하는 GR 신호전달과 관련된 전사인자 Redd1과 Klf15의 mRNA 발현을 Dex 처리군 대비 각각 약 45.8%, 28.2% 하향 조절하였다.
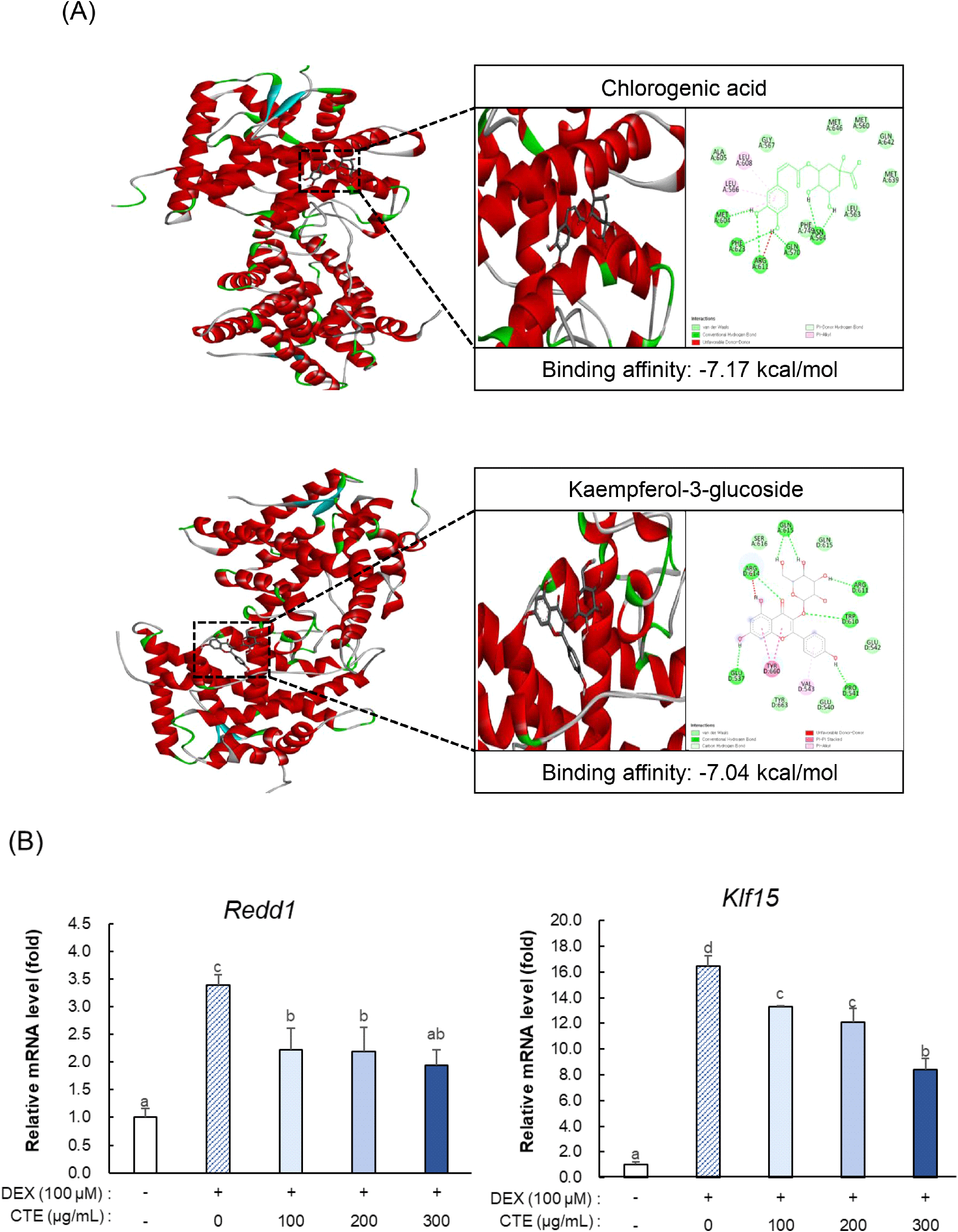
다양한 연구에서 천연물의 유효성분들이 GR과 결합함이 보고되었다. 본 연구의 molecular docking 분석 결과, CTE 내 주요 폴리페놀들이 GR의 활성 부위와 결합 가능성을 보였으며, 이는 이전 연구에서 보고된 다양한 천연물과 GR 간의 결합 경향과 유사하다(Sarmah, 2012). Kim 등(2025)은 molecular docking 분석을 통해 인삼과 홍삼의 주요 활성 사포닌 성분인 진세노사이드가 GR과 -9.4 kcal/mol의 높은 친화도로 결합함을 확인하였는데, 이는 Dex의 친화도인 -7.4 kcal/mol보다 높은 수치이다. 이를 통해 진세노사이드가 GR을 매개로 Dex 작용을 경쟁적으로 억제하여 근육 위축이 억제하는 기전을 제시하였다(Kim 등, 2025). GR은 근육 단백질 분해를 유도하는 주요 전사인자 및 관련 유전자들인 Klf15, Redd1, MuRF1, Atrogin-1 등의 발현을 조절하는 핵심 조절자이다(Permpoon 등, 2025). 따라서 Fig 4에서 확인된 CTE 처리에 의한 Redd1과 Klf15 발현 감소가, CTE에 함유된 주요 폴리페놀들이 GR에 작용하여 GR 매개 이화작용 유전자 전사를 억제함으로써 근위축 완화 효과를 나타낼 수 있음을 시사한다. 특히, molecular docking 분석 결과는 CTE 성분들이 Dex와 상호작용하는 GR의 활성 부위에 결합할 가능성을 제시하지만 이를 확인하기 위한 추가적인 실험이 필요하다. 결론적으로, Dex가 GR을 통해 유도하는 근육 단백질 분해 증가 및 단백질 합성 감소로 인해 발생하는 근위축은 CTE 내 유효성분들의 GR 상호작용을 통해 단백질 항상성이 회복되어 완화된다(Supplementary Fig. S1).
4. 요약
본 연구에서는 Dex로 유도된 근위축 C2C12 근관세포 모델에서 CTE의 근위축 억제 효과와 그 기전을 분석하고자 하였다. CTE는 DPPH 및 ABTS 라디칼 소거능과 TPC 및 TFC 값을 통해 우수한 항산화 효능을 보였다. HPLC 분석을 통해 chlorogenic acid, isoquercetin 및 kaempferol-3-glucoside가 CTE의 유효성분임을 확인하였다. C2C12 근관세포에 대한 세포 독성 평가에서 CTE는 최대 300 μg/mL에서도 세포 생존율을 저해하지 않았다. Dex로 유도된 근위축 모델에 CTE를 처리한 결과, Dex 처리군 대비 감소하였던 근관 직경과 MyHC 발현을 유의하게 증가시켰다. 또한, Dex 처리 시 감소했던 단백질 합성 관련 신호인 p-Akt와 p-mTOR 발현 역시 상향 조절하였으며, 반면에 Dex 처리에 의해 증가했던 단백질 분해 관련 지표인 MuRF1의 발현은 유의하게 억제되었다. Molecular docking 분석결과, CTE 내 주요 폴리페놀인 chlorogenic acid와 kaempferol-3-glucoside가 GR 활성 부위와 안정적으로 결합함을 확인하였다. CTE 처리 시 Redd1 및 Klf15 유전자 발현이 감소하는 결과는 폴리페놀과 GR 결합을 통한 GR 매개 이화작용 경로의 억제 기전을 분자수준에서 뒷받침한다. 종합적으로, 본 연구는 CTE가 항산화 효과 및 GR 경로와의 상호작용을 통한 단백질 항상성 조절 기전을 통해 Dex 유도 근위축을 효과적으로 완화할 수 있음을 확인하였다. 이는 CTE의 주요 폴리페놀들이 GR 경로를 조절하여 근육 단백질 항상성을 조절하는 핵심 기전임을 시사한다. 이러한 결과는 꾸지뽕나무잎을 활용한 근위축 예방 및 치료용 기능성 소재 개발의 과학적 근거를 마련하는 데 기여할 것으로 사료된다.