








1. Introduction
Skin color is influenced not only by genetic factors but also by several environmental stimuli, such as UV exposure, hormonal fluctuations, and inflammation (Snyman et al., 2024). Melanin plays a vital physiological role in protecting cell nuclei from UV-induced damage (Herraiz et al., 2021). However, excessive melanin production, or hyperpigmentation, can lead to aesthetic skin conditions such as melasma, freckles, and other pigmentation disorders (Moolla et al., 2022). Consequently, regulation of the melanogenic pathway has become a major focus of cosmetic and biomedical research aimed at developing effective skin-whitening strategies (Jo et al., 2024). Melanin biosynthesis comprises a complex cascade of biochemical reactions catalyzed by several key enzymes (D’Mello et al., 2016). Among these, tyrosinase serves as the rate-limiting enzyme in melanogenesis, catalyzing the oxidation of L-tyrosine to 4-Dihydroxy-L-phenylalanine ethyl ester (L-DOPA) and subsequently to dopaquinone (Yamazaki et al., 2025). Subsequent conversion steps leading to eumelanin intermediate formation are catalyzed by TRP-1 and TRP-2 (Wagatsuma et al., 2023). MITF serves as the principal transcriptional regulator of these enzymes, governing melanocyte differentiation, survival, and the expression of melanogenic genes (Chauhan et al., 2022). The transcriptional activity and expression of MITF are modulated by several upstream signaling cascades, notably the MAPK, PKA, and CREB pathways. The MAPK pathway, which plays a pivotal role in cell proliferation, differentiation, and stress responses, is subdivided into three branches: ERK, JNK, and p38 MAPK (Cánovas et al., 2021). Sequential phosphorylation events within these MAPK cascades lead to kinase activation, nuclear translocation, and subsequent phosphorylation of transcription factors such as MITF, ultimately altering the expression of genes related to melanogenesis (Kim et al., 2024b). Extracellular ligands such as α-MSH bind to melanocortin 1 receptor on the melanocyte surface, activating adenylate cyclase and increasing intracellular cAMP levels (Arora et al., 2021). Elevated cAMP levels activate PKA, which phosphorylates CREB into its active form (Ko and Lee, 2021). Activated CREB binds to the promoter region of MITF, enhancing its transcription and subsequently upregulating the expression of enzymes involved in melanin biosynthesis (Zhou et al., 2021).
A member of the Zingiberaceae family, Kaempferia parviflora is a perennial herbaceous plant widely recognized for its medicinal properties. In Thailand, it is commonly known as “Krachai Dum” or “Thai ginseng” and naturally grows across Southeast Asia. Its rhizome displays a characteristic purple-to-black coloration, which gives rise to the name “black ginger.” It is recognized as one of Thailand’s five key medicinal plants (Tan et al., 2024). Traditionally, black ginger has been used as a folk remedy for treating various ailments, including inflammation, allergy, ulcers, osteoarthritis, and gout (Chen et al., 2018). Recent pharmacological studies have demonstrated that black ginger extracts possess diverse biological activities, such as antioxidants, anti-inflammatory, anti-obesity, anticancer, cardioprotective, neuroprotective, and antimicrobial effects (Pham and Hong, 2025). These biological activities are primarily attributed to the polymethoxyflavones (PMFs) abundantly present in their rhizome. Notably, these compounds suppress the hallmarks of aging, including ROS generation and the SASP, thereby preserving the integrity of human dermal fibroblasts (Huo et al., 2023). A recent study demonstrated that black ginger extract and its methoxyflavones, particularly 5,7,4′-trimethoxyflavone, significantly ameliorated memory deficits in scopolamine-induced mice by inhibiting cholinesterase activity and preventing amyloid-β aggregation (Takomthong et al., 2025). They promote tropocollagen synthesis and restore the expression of ECM components, including procollagen, fibrillin-1, and hyaluronic acid, improving skin aging (Klinngam et al., 2022).
Black ginger is thus an important medicinal plant traditionally used for treating several diseases, with emerging evidence supporting its diverse therapeutic potential. Additionally, since black ginger has long been consumed as a traditional food, it offers the advantage of safety for dietary intake, making it a promising candidate for inner beauty. However, there is currently a paucity of literature regarding its efficacy as a melanin synthesis inhibitor. This research was designed to assess the efficacy of black ginger as a suppressor of melanin production for functional ingredients in inner beauty.
2. Materials and methods
K. parviflora (black ginger) was obtained from SNS Company (Bangkok, Thailand). The raw materials were cleansed and dehydrated at 50°C for a period of 72 h. Following dehydration, the black ginger was subjected to sonication using 70% ethanol for 24 h. The solution was purified via filtration, and the solvent was subsequently eliminated through vacuum evaporation. The extract was stored at −20°C to maintain stability prior to analysis.
Electron donating ability (EDA) was determined using a modified Blois method. (Blois, 1958). Extracts were mixed with 80 μM ethanolic DPPH, incubated for 15 min in the dark, and absorbance was measured at 517 nm. ABTS radical scavenging activity followed the method from Re et al (1999). ABTS radicals, generated by reacting 7 mM ABTS with 2.45 mM potassium persulfate for 24 h, were diluted with ethanol. Extracts were mixed with ABTS reagent (1:1) and absorbance was read at 700 nm after 1 min. The radical scavenging activity for both assays was calculated using the following equation:
Mouse melanoma B16F10 cell line was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, Grand Island, NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 IU/mL penicillin, and 100 μg/mL streptomycin. All cultures were kept at 37°C with 5% CO2.
B16F10 cells were plated at 1×104 cells/well and exposed to KEE (0-1,000 μg/mL) for 24 h. Cell viability was determined via the MTT assay. Briefly, MTT solution was added for 4 h, and the insoluble formazan was solubilized with DMSO. The absorbance was recorded at 540 nm to quantify viable cells.
Melanin levels in B16F10 cells were measured using a modified protocol from Kang and Lee (2025). Cells (1×106 cells/well) were co-treated with α-MSH and KEE for 24 h. The cells were lysed and the pellets were solubilized in a 1 N NaOH/10% DMSO solution at 90°C. To account for differences in cell density, melanin content was normalized to the total protein content determined via a bicinchoninic acid method (Thermo Fisher Scientific, MA, USA). Absorbance was measured at 490 nm.
B16F10 cells were exposed to α-MSH (100 nM) with or without KEE (5, 10, and 50 μg/mL) for 24 h. Total RNA was extracted using TRIzol reagent (Invitrogen, CA, USA), and cDNA was synthesized from 5× Green GoTaq Flexi Buffer, 1 μg RNA using 1 mM dNTPs, oligo (dT) primers and Taq DNA polymerase (Promega, WI, USA). GAPDH served as the internal control. The following primer sequences were used for amplification of the target genes: GAPDH (reverse) AGCCTTCTCCATGGTGGTGAAGAC, (forward) CGGAGT CAACGGATTTGGTCGTAT; MITF (reverse) TAGCTCCTT AATGCGGTCGT, (forward) AGCGTGTATTTTCCCCACAG; Tyrosinase (reverse) GCCATGACCAGGATGAC, (forward) GACGGTCACTGCAGACTTTG.
B16F10 cells were cultured for 24 h, treated with KEE, and subsequently exposed to α-MSH. The cells were washed with PBS and lysed using RIPA buffer supplemented with inhibitors. Cell lysates were clarified by centrifugation and protein levels were measured using a bicinchoninic acid protein assay. Equivalent amounts of protein were resolved by SDS-PAGE. Subsequently the separated proteins were electrotransferred onto membranes. They were incubated in 5% non-fat milk for 1 h to block nonspecific binding, then probed overnight at 4°C with primary antibodies. After rinsing in TBST, membranes were subjected to appropriate secondary antibodies for 1 h. Immunoreactive signals were detected using enhanced chemiluminescence reagents and analyzed by densitometry with the imaging system.
All experiments were conducted in triplicate or more, independently. Results are reported as mean±SD. Statistical analysis was performed using SPSS version 23.0 (IBM Corp., Armonk, NY, USA). Statistical significance was determined using one-way analysis of variance followed by Duncan’s multiple range test at a significance level of p<0.05.
3. Results and discussion
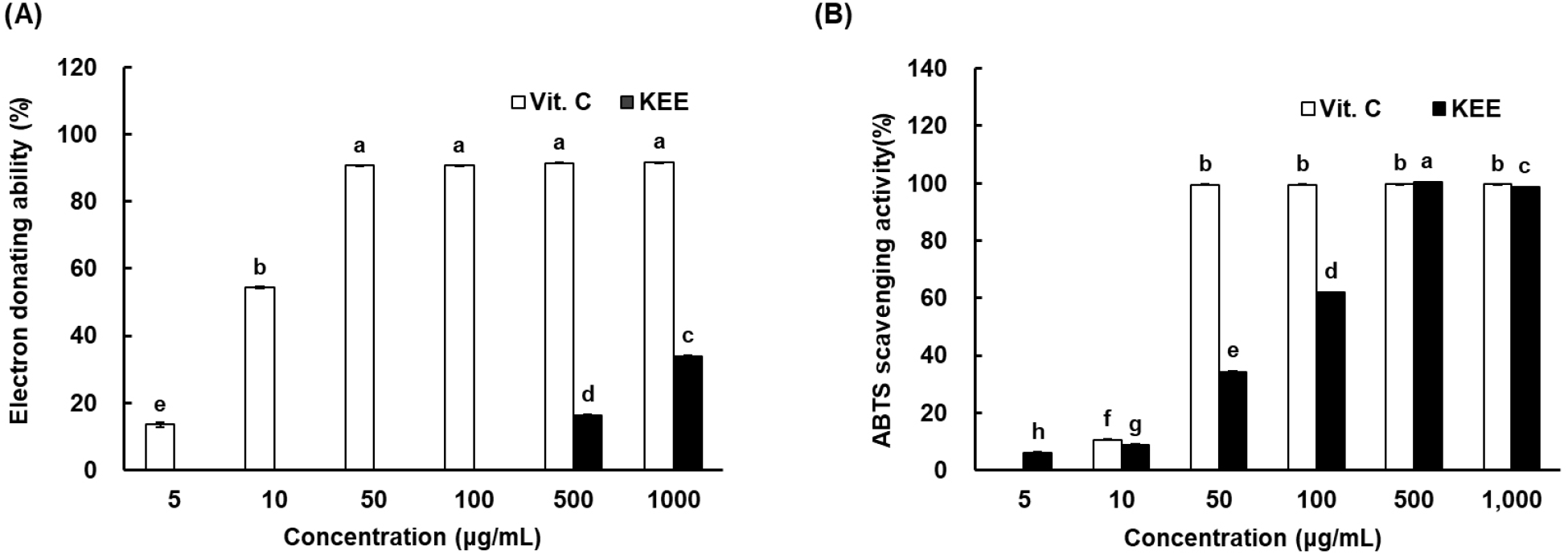
The antioxidant activity of K. parviflora extract (KEE) was assessed via DPPH and ABTS radical scavenging assays. DPPH reacts with hydrogen or electron donors, changing color from purple to yellow upon reduction, while ABTS+, a stable cationic radical, measures both hydrophilic and lipophilic antioxidant activities. Because DPPH generates an anionic radical and ABTS a cationic one, differences in their reactivity may lead to differences in antioxidant responses. The DPPH radical scavenging capacity of KEE was 16.13% at 500 μg/mL and 33.78% at 1,000 μg/mL (Fig. 1). However, the ABTS+ radical scavenging activity reached 99.24% and 98.60% at the same concentrations, respectively. Notably, the ABTS+ scavenging capacity of KEE was comparable to that of vitamin C, indicating strong radical scavenging potential. Traditionally, hydroquinone, vitamin C, kojic acid, and their derivatives have been widely used as depigmentation agents. However, hydroquinone has been associated with adverse effects such as skin irritation, exogenous ochronosis, and contact dermatitis (Zhu et al., 2008). Consequently, natural products rich in phytochemicals, such as flavonoids, phenolic acids, and their derivatives, have demonstrated diverse biological activities (Chiocchio et al., 2018). These results demonstrated that KEE possesses significant antioxidant activity in vitro, prompting an investigation into its effects on melanogenesis and the underlying molecular mechanisms.
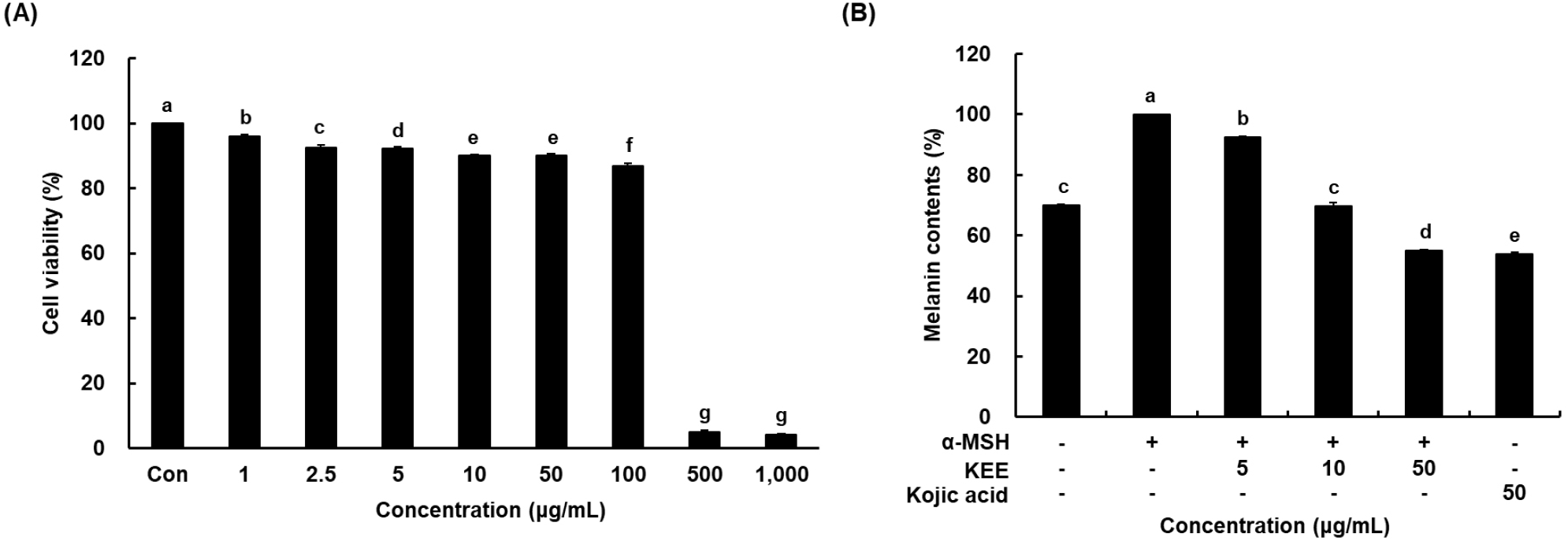
The cytotoxicity of KEE in melanoma cells was determined by an MTT method. Treatment with KEE at concentrations up to 1,000 μg/mL exhibited no apparent cytotoxicity below 100 μg/mL, and cell viability remained above 90% (Fig. 2A). Therefore, the concentrations of 5, 10, and 50 μg/mL were selected for subsequent assays of melanin production in α-MSH-stimuli. To evaluate the effect of KEE on melanogenesis, B16F10 cells were pretreated with 100 nM α-MSH for 24 h to induce melanogenesis, followed by incubation with various concentrations of KEE. Melanin production was substantially upregulated in response to α-MSH stimulation relative to the control. Treatment with KEE dose-dependently decreased intracellular melanin levels, suggesting that KEE exerts an inhibitory effect on melanin production without affecting cell viability (Fig. 2B). Notably, KEE exhibited only a slight increase in direct tyrosinase inhibitory activity compared to the control group. Melanin biosynthesis occurs in melanocytes, specialized cells containing tyrosinase and related enzymes catalyzing the conversion of precursors into melanin. Tyrosinase plays a key role in melanin synthesis by catalyzing the initial steps tyrosine oxidation leading to melanin formation (Horibe et al., 2013). The anti-melanogenic efficacy of K. parviflora is primarily attributed to its abundance of bioactive methoxyflavones, including 5,7-dimethoxyflavone, 3,5,7,3’,4’-pentamethoxyflavone, and 5,7,4’-trimethoxyflavone (Chaisuwan et al., 2022). Previous studies on structure-activity relationships (SARs) have demonstrated that the activity of these compounds depends critically on the specific positions of methoxy groups. Notably, potent inhibitory effects are linked to methoxy substitutions at the C-5 and C-4’ positions, whereas a methoxy group at the C-3 position tends to diminish bioactivity (Huo et al., 2023). Considering these structural determinants, the significant anti-melanogenic activity observed in our study suggests that the extract acts through the synergistic potency of these key methoxyflavones, particularly those lacking C-3 substitution. Collectively, these findings indicate that KEE likely inhibits melanogenesis by regulating upstream signaling pathways and transcription factors rather than by directly inhibiting tyrosinase activity.
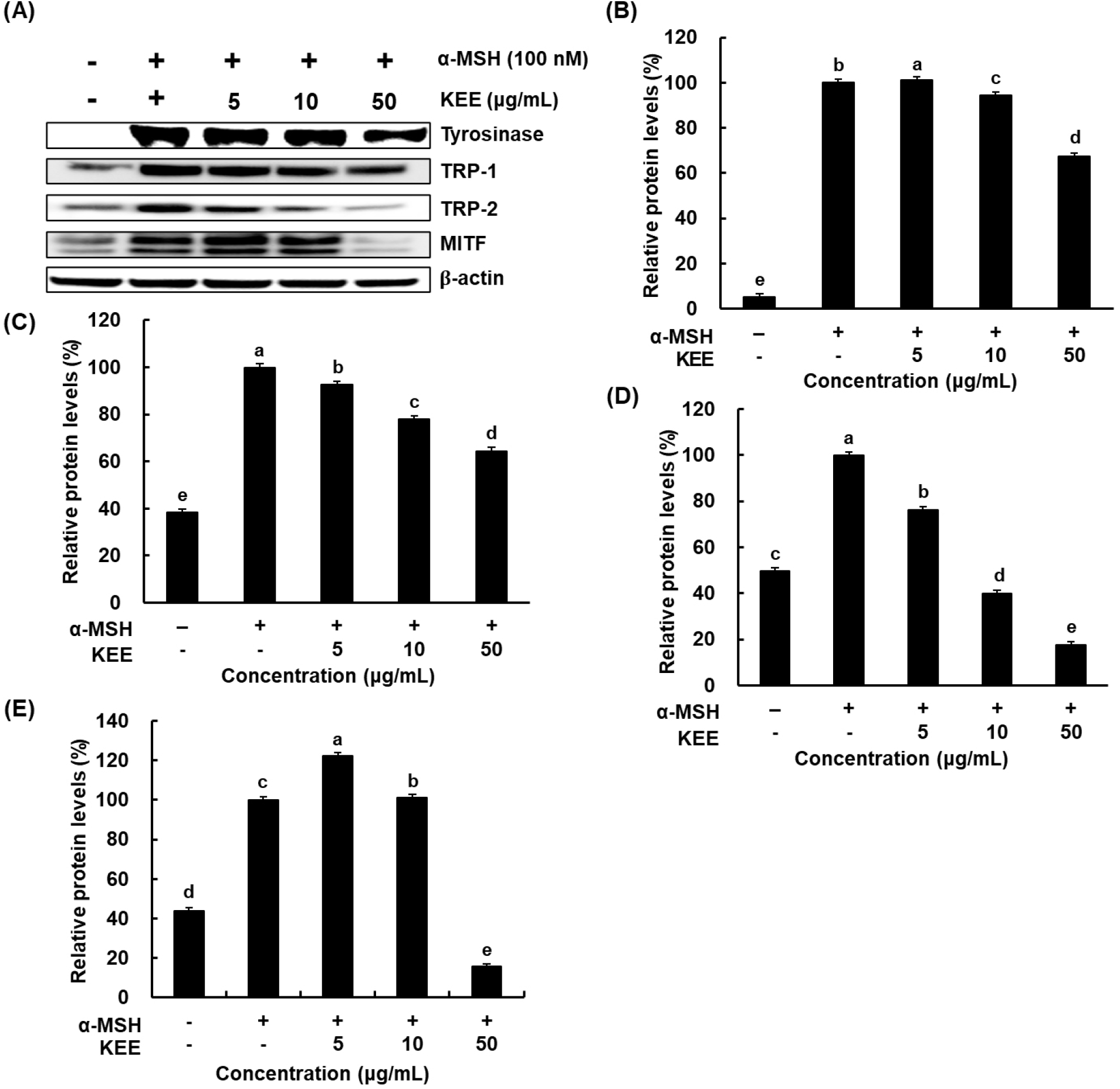
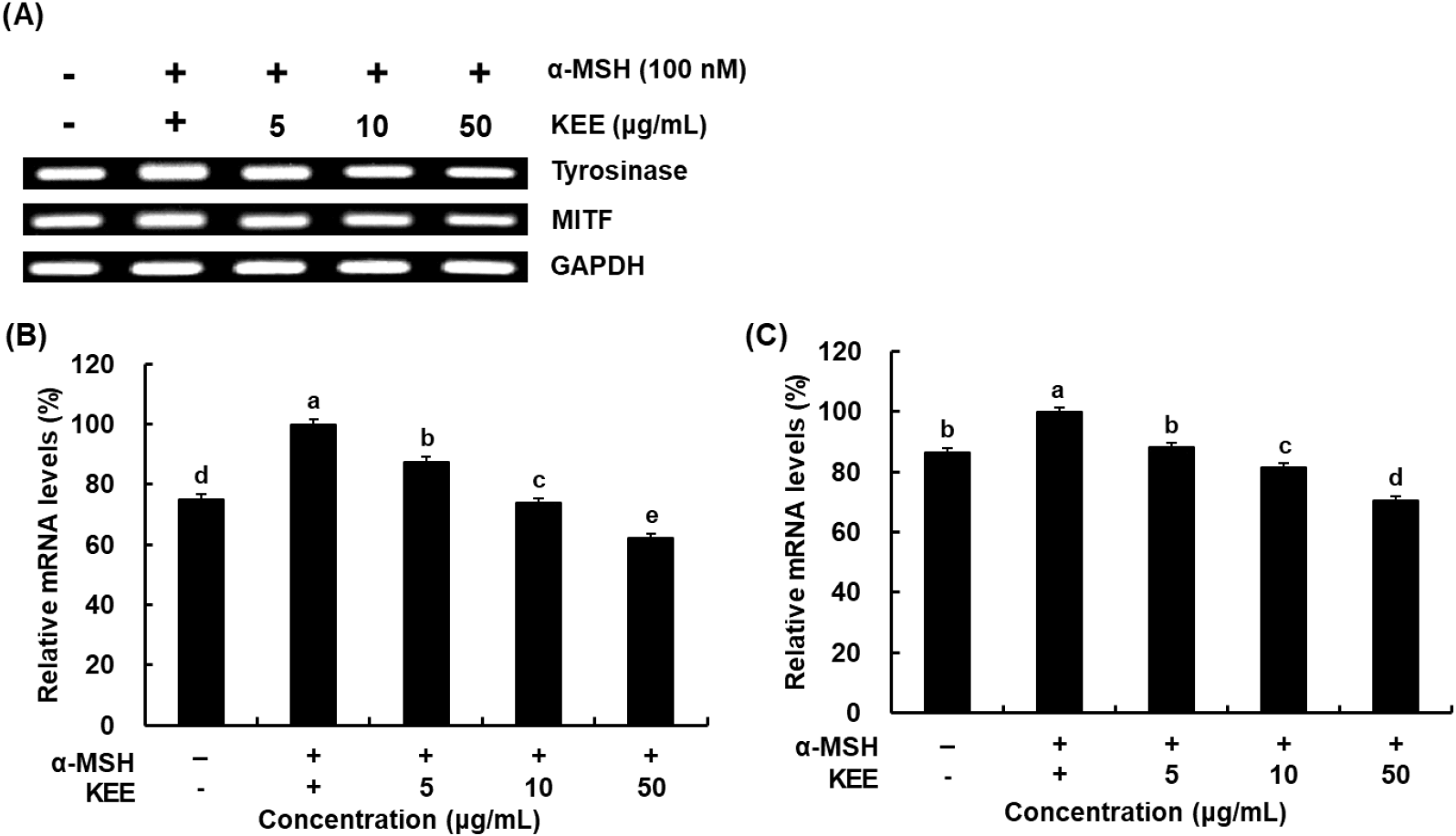
Melanin synthesis is initiated and regulated by several signaling pathways and transcription factors, including those involved in tyrosinase activation. In addition to melanogenic genes are localized in melanosomes and catalyze key reactions in eumelanin biosynthesis. It has been reported that α-MSH induces the levels of tyrosinase during melanogenesis (Briganti et al., 2003). MITF acts as a transcriptional regulator of melanogenic enzymes such as tyrosinase, TRP-1, and TRP-2 (Kim et al., 2021). Western blotting was employed to elucidate whether KEE modulates the expression levels of melanogenic enzymes in cells stimulated with α-MSH. The expressions of TRP-1, TRP-2, and tyrosinase were decreased in KEE-treated cells compared with the α-MSH-stimulated group (Fig. 3). Notably, treatment with 50 μg/mL KEE reduced these protein levels by more than 30%. Given that KEE inhibited tyrosinase-related protein expression, its effect on MITF, a key transcription factor involved in melanogenesis was therefore further examined. The expression of MITF was markedly increased upon α-MSH stimulation but significantly downregulated by KEE. Furthermore, KEE reduced both tyrosinase and MITF mRNA levels, consistent with the observed protein expression data (Fig. 4). These findings indicate that KEE inhibits melanogenesis by suppressing MITF expression and subsequently attenuating the expression of MITF-regulated melanogenic enzymes. Similarly, Momordica cochinchinensis extract has been reported to suppress MITF and its downstream melanogenic enzymes, indicating that MITF regulation is a common mechanism among natural anti-melanogenic agents (Yoo et al., 2021). Consistent with these findings, KEE exhibited inhibitory effects on melanin production comparable to those of kojic acid, suggesting that it possesses potent anti-melanogenic activity similar to other plant-derived compounds.
Prior investigations have verified that MITF expression is regulated by several signaling pathways involved in melanogenesis. Among these, kinases that act on the CREB influence MITF transcription. α-MSH promotes melanogenesis by elevating intracellular cAMP levels and enhancing CREB phosphorylation, which subsequently activates cAMP-dependent PKA through cAMP-mediated signaling (Kim et al., 2024a). Furthermore, the cAMP/PKA/CREB signaling axis constitutes another critical regulatory pathway in melanin synthesis. This signaling cascade responds to extracellular stimuli and modulates MITF activity through regulation at the transcriptional and post-translational stages, thereby controlling the overall melanogenic response (Yoo et al., 2021). The inhibitory potential of KEE on the PKA/CREB axis was investigated by assessing the activation status of these signaling proteins. CREB phosphorylation was significantly reduced in cells treated with both α-MSH and KEE compared with those treated with α-MSH alone, demonstrated that KEE treatment inhibited the phosphorylation of PKA at threonine 197 and CREB at serine 133 in α-MSH-stimulated cells (Fig. 5). Similarly, KEE treatment decreased PKA phosphorylation. Our data suggest that the inhibitory mechanism of KEE on melanin synthesis is mediated via the attenuation of the cAMP/PKA/CREB cascade.
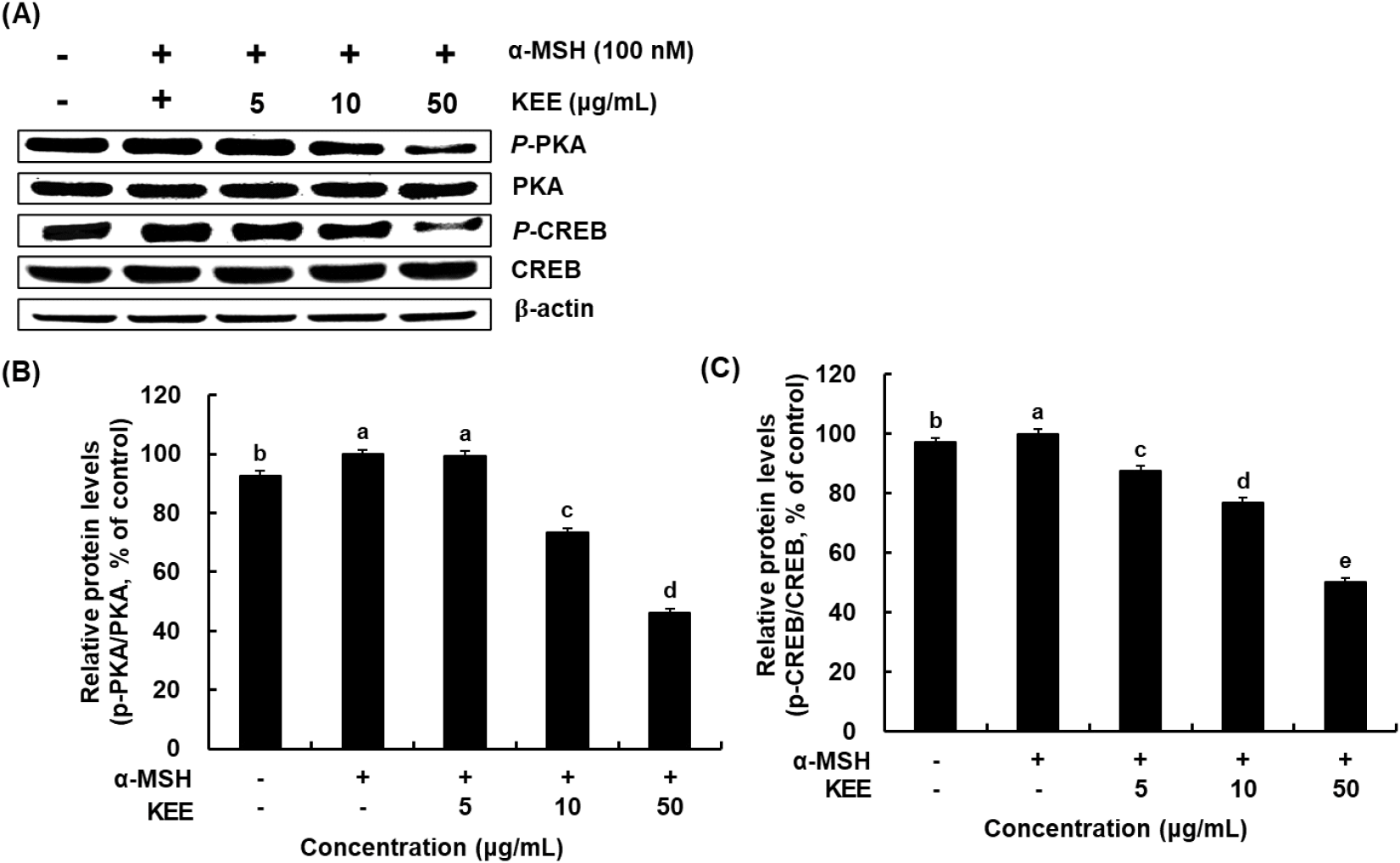
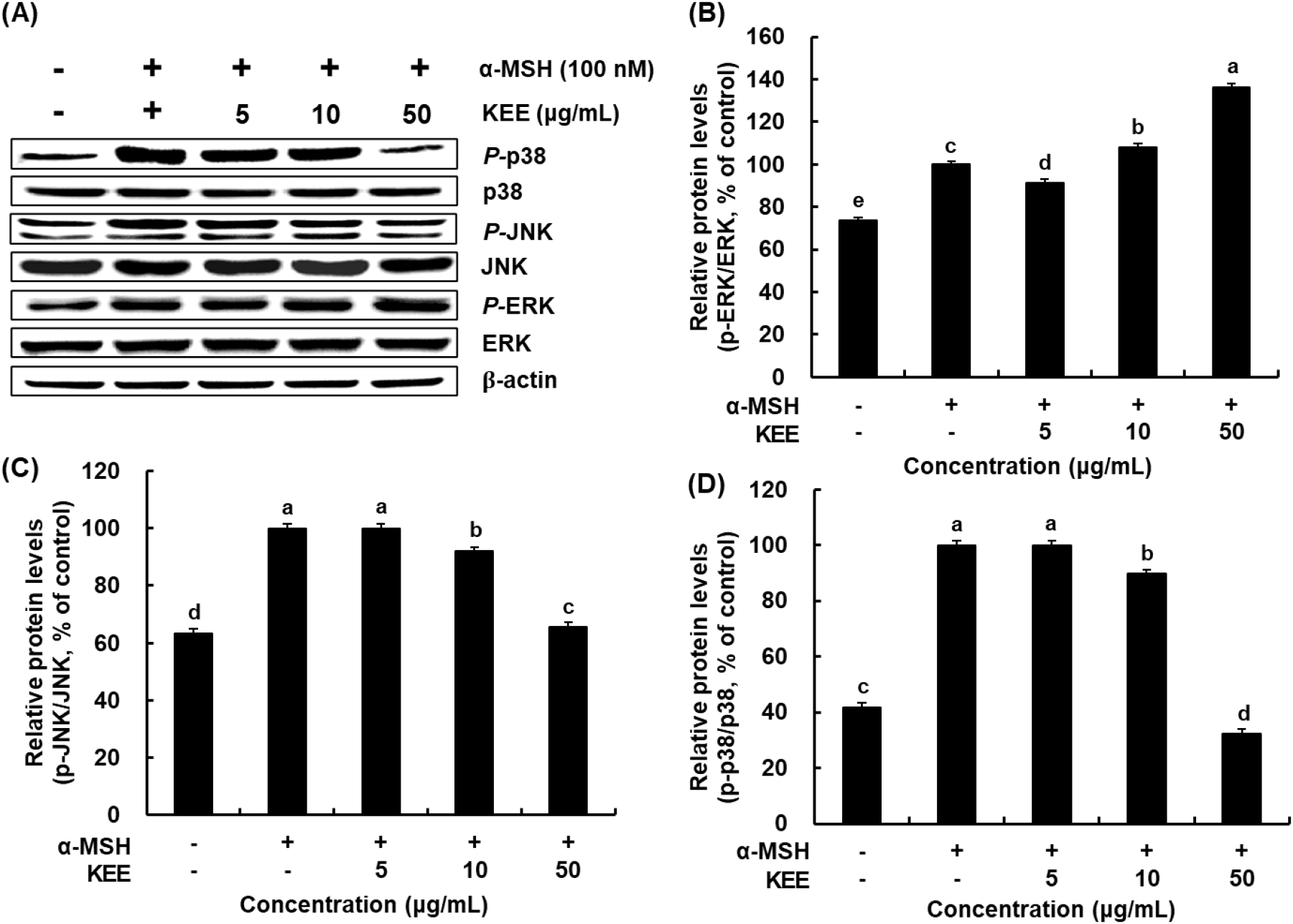
The MAPK pathway plays a major regulatory role in melanogenesis by modulating MITF expression. Phosphorylation events within the MAPK signaling cascade influence the activity of melanogenic enzymes such as tyrosinase, thereby preventing excessive melanin synthesis (Zhou et al., 2022). We investigated the modulatory influence of KEE on the MAPK signaling pathways by analyzing the phosphorylation of p38, ERK, and JNK. KEE treatment markedly reduced the α-MSH-induced phosphorylation of p38 and JNK, whereas ERK phosphorylation was slightly increased by KEE at a concentration of 50 μg/mL (Fig. 6). These signaling cascades converge on MITF to stimulate the synthesis of melanogenic enzymes and subsequent pigment production (Kim et al., 2024a). α-MSH stimulation increases cAMP levels and induces PKA phosphorylation, which regulates the downstream CREB. CREB phosphorylation may also occur via MAPK, which affect MITF expression in melanocytes (D’Mello et al., 2016). Collectively, KEE suppresses melanogenesis by regulating PKA/CREB/MAPK signaling pathway in α-MSH-stimulated melanocytes, thereby demonstrating its potential as a natural depigmentation compound.
4. Conclusions
K. parviflora (black ginger) extract exhibits potent anti-melanogenic activity in α-MSH-stimulated B16F10 melanocytes. KEE significantly reduced melanin production without directly inhibiting tyrosinase activity, indicating that its depigmenting effect is primarily mediated through upstream transcriptional regulation. KEE treatment exerted suppressed expression of the master regulator MITF. This reduction was accompanied by a concomitant decrease in the expression levels of the key melanogenic enzymes. Also, KEE suppressed melanogenesis by inhibiting the cAMP/PKA/CREB signaling cascade and modulating MAPK-related pathways, culminating in reduced MITF expression and melanin production. KEE exerts anti-melanogenic effects through regulation of the PKA/CREB/MAPK signaling pathway. Therefore, KEE could be developed as a dual function agent for both cosmetic ingredients and dietary supplements for inner beauty to suppress melanogenesis.