








1. Introduction
Bamboo shoots have been valued as nutritious and functional foods for centuries, particularly in Asia, where they form an integral component of traditional cuisine. These young, edible bamboo culms are characterized by their low fat content, high dietary fiber, and rich mineral profile. This combination makes them both a culinary staple and a source of important nutrients (Basumatary et al., 2017). While bamboo shoots have traditionally been harvested by indigenous and rural communities, their popularity has expanded significantly into urban markets and the global culinary scene. This expansion stems from increasing recognition of their distinctive flavor profile and health-promoting properties.
The nutritional composition of bamboo shoots reveals a diverse array of macro- and micronutrients that contribute to their status as a functional food (Nongdam and Tikendra, 2014). Typically, bamboo shoots contain approximately 88.8% water, over 3.9% protein. Their amino acid profile is comprehensive and rivals many common vegetables, such as cabbage, carrots, and onions (Chongtham et al., 2021; Satya et al., 2010). Research has identified 17 amino acids in bamboo shoots. Tyrosine is typically the most abundant, while lysine is commonly the limiting amino acid across most bamboo species (Nongdam and Tikendra, 2014).
Beyond their protein content, bamboo shoots serve as excellent sources of essential micronutrients. These include chromium, zinc, manganese, and iron, which play crucial roles in various physiological functions (Oriwo et al., 2022). Their substantial dietary fiber content enhances digestive health. Plant sterols contribute to cardiovascular wellbeing through cholesterol regulation (Nongdam and Tikendra, 2014). Additionally, bamboo shoots contain phenolic compounds with documented antioxidant properties. These compounds can mitigate oxidative stress and potentially reduce the risk of chronic disease (Park and Jhon, 2010). This comprehensive nutritional and nutraceutical profile positions bamboo shoots as valuable components of health-conscious diets.
From a practical harvesting perspective, the culinary value of bamboo shoots is influenced by the yield of their edible portion. This typically ranges from 29% to 33% depending on species and harvesting practices (Sood et al., 2017). Some investigations have reported that up to 70% of harvested shoot material may be discarded as waste during the processing stage. This waste consists primarily of outer sheaths and basal sections, highlighting opportunities for improved utilization efficiency (Lin et al., 2018). In Thailand, several bamboo species are commercially harvested for their high-quality edible shoots. These include Dendrocalamus asper, Dendrocalamus giganteus, Dendrocalamus merrillianus, Bambusa tulda, Pseudoxytenanthera albociliata, and Thyrsostachys siamensis. However, a significant knowledge gap remains regarding the nutritional and functional properties of less-studied species. Bambusa beecheyana and Dendrocalamus latiflorus, in particular, show commercial potential in the region but remain understudied.
Traditional preparation methods for bamboo shoots typically involve boiling to remove naturally occurring cyanogenic glycosides. These compounds impart bitterness and may present toxicity concerns if consumed raw. This thermal processing not only improves palatability but may significantly alter the nutritional composition and bioactive compound profile of the shoots. Understanding these processing-induced changes is essential for accurately assessing their nutritional value in typical consumption patterns.
The present study addresses these knowledge gaps through a comprehensive nutritional and functional evaluation of two commercially promising yet understudied bamboo species: Bambusa beecheyana and Dendrocalamus latiflorus. To our knowledge, this is the first comparative investigation examining proximate composition, amino acid profiles, ash content, vitamin levels, antioxidant activities, and bioactive compounds in both fresh and boiled shoot samples from these species. By characterizing the impact of traditional boiling on the nutritional properties of foods, this study provides critical data for food composition databases and dietary recommendations. These findings will support the development of value-added bamboo-based food products that maximize nutritional benefits while maintaining consumer acceptability. Given the growing global interest in plant-based, sustainable food sources, elucidating the functional food potential of this bamboo species is particularly relevant for diversifying nutrient-rich options in both traditional and modern diets worldwide.
2. Materials and methods
The juvenile bamboo shoots of Bambusa beecheyana and Dendrocalamus latiflorus were obtained from local markets in Phayao, Thailand. The shoots were peeled, rinsed, and sliced longitudinally into strips with a thickness of 2-3 mm. The slices were weighed to 1,000 g and divided into two equal portions: one for the fresh treatment and the other for the boiling treatment. For boiling, the 500 g of slices were boiled in preheated distilled water (95-98°C) at a sample-to-water ratio of 1:10 (w/w) for 10 minutes to reduce cyanogenic glycosides, particularly taxiphyllin, which can release toxic hydrogen cyanide (HCN) upon hydrolysis (Pandey and Ojha, 2014). After heating, the slices were removed, cooled to room temperature, packed in polyethylene bags, and stored at 4°C until subsequent analyses.
The proximate composition, including moisture, protein, crude fat, ash, crude fiber, and N-free extract, was analyzed according to the official methods of the Association of Official Analytical Chemists (AOAC, 2000). Moisture content was determined by oven-drying at 130°C to constant weight (AOAC 925.10). Ash content was obtained by incineration in a muffle furnace at 550°C (AOAC 923.03). Crude fat was quantified by Soxhlet extraction using petroleum ether as the solvent (AOAC 920.39). Crude fiber was measured by sequential acid and alkali digestion (AOAC 962.09). Protein content was analyzed by the Kjeldahl method (AOAC 984.13), with nitrogen values converted to protein using a conversion factor of 6.25. N-free extract content was calculated by difference according to the following equation:
All experiments were conducted with three biological replicates (n=3), each analyzed in duplicate. All values were expressed on a dry weight basis, except for moisture, which was reported on a wet weight basis.
Dietary fiber content was determined by the enzymatic-gravimetric method (AOAC 985.29). Samples were defatted prior to analysis and then subjected to sequential enzymatic digestion to simulate gastrointestinal conditions. Heat-stable α-amylase was used to hydrolyze starch, protease was applied to degrade protein, and amyloglucosidase was employed to remove residual starch. The undigested fraction was precipitated with ethanol, filtered, dried, and weighed to determine the total dietary fiber content. Corrections were applied to the protein and ash content in the residue to ensure accuracy. All analyses were performed in duplicate.
Amino acid profiles of bamboo shoots were analyzed using an amino acid analyzer (Model 8500L, Hitachi High-Technologies, Tokyo, Japan) equipped with an ion-exchange column (Hitachi amino acid column, 4.6×60 mm). Analyses were performed via postcolumn ninhydrin derivatization following the method of Yongsawatdigul et al. (2007) with minor modifications. For most amino acids, samples were hydrolyzed with 6 N hydrochloric acid (HCl) at 110°C for 24 h under a nitrogen atmosphere to prevent oxidation. The hydrolysates were filtered, evaporated to dryness, and reconstituted in sodium citrate buffer. Amino acids were separated based on ionic characteristics using the ion-exchange column and subsequently reacted with the Ninhydrin reagent. Detection was carried out photometrically at 570 nm for most amino acids and 440 nm for specific residues. Quantification was achieved by comparing peak areas with those of known amino acid standards. Tryptophan was determined separately by alkaline hydrolysis with 4.2 N sodium hydroxide (NaOH) at 110°C for 20 h, followed by neutralization, filtration, and subsequent analysis using the same amino acid analyzer system.
Nutritional quality indices were also calculated. The amino acid score (AAS) and essential amino acid index (EAAI) were derived according to Machado et al. (2020) The AAS was determined using reference amino acid requirements established by FAO/WHO/UNU (2007). The percentage contribution to daily amino acid requirements was estimated based on reference intake values for fresh vegetables specified in Thailand’s nutrition labeling regulations (Ministry of Public Health, 2023).
The phytosterol content was determined by gas chromatography (GC) using the method of Lagarda et al. (2006) with minor modifications. Approximately 250 g of bamboo shoots was used for the analysis. Lipids were first extracted with petroleum ether and then subjected to saponification with ethanolic KOH at room temperature. To release conjugated phytosterols, acid hydrolysis was subsequently performed, after which the unsaponifiable fraction was extracted. Phytosterols were separated and quantified using a gas chromatograph (Agilent 7890B GC System, Agilent Technologies, USA) equipped with a flame ionization detector (FID) and an HP-5MS capillary column (30 m×0.25 mm i.d., 0.25 μm film thickness). The injector and detector temperatures were set at 250°C and 280°C, respectively. The oven temperature program was as follows: initial temperature, 40°C (held for 3 min), increased at 5°C/min to 200°C, followed by a 10°C/min increase to 250°C, and held for 5 min. Helium was used as the carrier gas at a flow rate of 1.0 mL/min, and 1 μL of sample was injected in splitless mode. The following phytosterols were identified and quantified: campesterol, stigmasterol, β-sitosterol, stigmastanol, and brassicasterol.
Total phenolic content was determined using the Folin-Ciocalteu colorimetric method (Lee et al., 2020). Samples were extracted with 80% methanol, and 0.1 mL of the extract was mixed with 0.5 mL of distilled water, followed by 0.1 mL of Folin-Ciocalteu reagent. Subsequently, 1 mL of 7% sodium carbonate (Na2CO3) solution was added, and the mixture was thoroughly mixed and incubated in the dark at room temperature for 90 min. Absorbance was measured at 760 nm using a UV-Vis spectrophotometer (UV-1800, Shimadzu Corporation, Japan). Results were expressed as milligrams of gallic acid equivalents per 100 g dry weight (mg GAE/100 g db).
The antioxidant capacity of bamboo shoots was evaluated using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging assay (Prommajak et al., 2014). Briefly, 0.1 mL of the methanolic extract was mixed with 5 mL of 0.1 mmol/L DPPH solution prepared in methanol. The mixture was vortexed for 1 min and incubated in the dark at room temperature for 20 min. Absorbance was then measured at 517 nm using a UV-Vis spectrophotometer. Antioxidant activity was expressed as milligrams of ascorbic acid equivalent per 100 g dry weight (mg AAE/100 g db).
All analyses were performed in triplicate, and the results were expressed as mean±SD. Differences among treatments were evaluated using analysis of variance (ANOVA), and significant differences were determined by Duncan’s multiple range test at p<0.05. Statistical analyses were conducted using Minitab version 18 (Minitab LLC, State College, PA, USA). Data visualization, including box-whisker plots and principal component analysis (PCA), was performed using R software version 4.2.2 (R Core Team, 2022).
3. Results and discussion
The proximate composition of fresh and boiled bamboo shoots from B. beecheyana and D. latiflorus is presented in Table 1. The moisture content of both species ranged from 92% to 93%, which is within the previously reported range for bamboo shoots (78-94%), depending on the species (Wang et al., 2020). Boiling significantly increased the moisture content, likely due to water absorption during thermal processing. This observation is consistent with earlier studies on Phyllostachys praecox, Bambusa burmanica Gamble, and Thysostachys siamensis Gamble, whereas moist-heat cooking methods such as boiling and steaming led to increased water content, while dry-heat treatments such as stir-frying resulted in moisture loss (Wang et al., 2020; Zhang et al., 2011).
B. beecheyana contained significantly higher protein and lower fat levels compared with D. latiflorus (p<0.05), whereas ash content did not differ significantly between the two species (Table 1). However, boiling significantly increased fat content in both species (128.9% in B. beecheyana, 228.8% in D. latiflorus, p<0.05). In contrast, the protein content in D. latiflorus decreased markedly by 62.3%, while that of B. beecheyana exhibited negligible change (0.26% loss). Ash content also declined substantially in both species, with a 40.6% reduction observed, indicating notable mineral leaching into the boiling water. Similar trends have been observed in other vegetables. The increase in fat content following boiling may be attributed to the relative concentration effect, resulting from the leaching of water-soluble components, and to improved lipid extractability due to cell wall disruption during heat treatment (Tenyang et al., 2017). In contrast, several studies have reported notable protein losses after boiling in various vegetables, such as red pepper and lotus rhizome, likely due to protein denaturation and solubilization (Chen and Mohamad, 2020; Hwang et al., 2012). The reduction in protein is mainly attributed to the leaching of nitrogenous compounds into the cooking medium, as well as thermal denaturation of heat-sensitive proteins. Likewise, ash content, which serves as an indicator of mineral levels, decreased significantly after the boiling process. A comparable reduction was reported in lotus rhizome, where boiling caused a decline in ash content due to the solubilization and leaching of water-soluble minerals (Chen and Mohamad, 2020). Additionally, studies on traditional Malawian vegetables have demonstrated overall nutrient losses, including minerals, after boiling (Issa et al., 2019).
The total dietary fiber (TDF) content of bamboo shoots in this study ranged from 38.16 to 49.44 g/100 g db, while crude fiber values were lower, ranging from 22.34 to 38.01 g/100 g db (Table 1). These results are consistent with reported ranges for Bambusa species (30.23-35.15 g/100 g db) and Dendrocalamus species (8.88-63.62 g/100 g db) (Wang et al., 2020). Because moisture content in plant materials varies with species, cultivation conditions, and postharvest handling, expressing proximate values on a dry weight basis provides a more accurate representation of nutritional composition. Notably, the TDF content of B. beecheyana and D. latiflorus in this study was at the upper end of the interquartile range reported for these genera (Fig. 1).
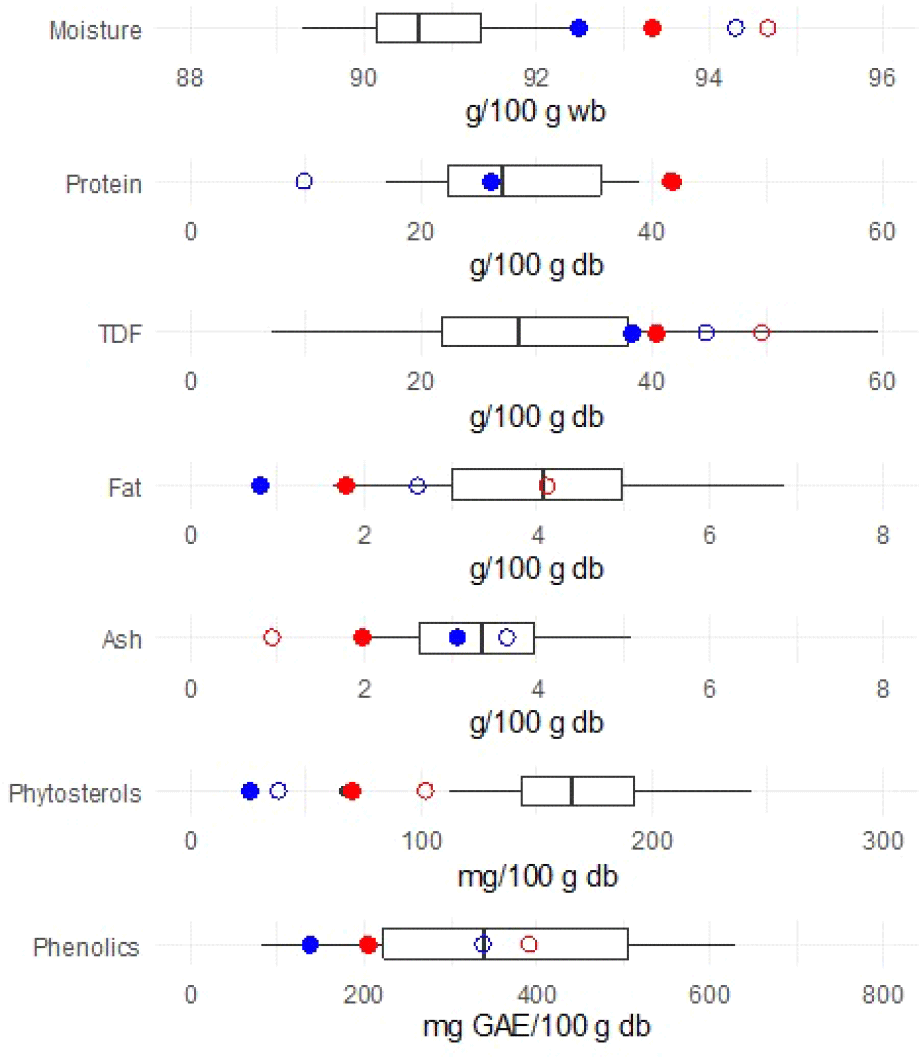
TDF consists of soluble dietary fiber (SDF) and insoluble dietary fiber (IDF), both of which are important for human health. IDF is typically the predominant fraction in bamboo shoots, contributing to gut motility and stool bulk, whereas SDF plays a metabolic role by modulating blood glucose and cholesterol levels (Chailangka et al., 2024). The current study showed a similar pattern, where crude fiber, primarily composed of IDF, constituted a major portion of TDF.
Boiling significantly increased both TDF and crude fiber contents in B. beecheyana and D. latiflorus (p<0.05; Table 1). A similar increase in crude fiber has been reported in lotus rhizomes after steaming and boiling (Chen and Mohamad, 2020). This apparent increase is unlikely to reflect structural modifications of dietary fiber, since cellulose, hemicellulose, and lignin are relatively stable at boiling temperatures. Instead, it may be explained by the leaching of soluble components such as proteins and minerals into the cooking medium, thereby increasing the relative proportion of fiber in the dry matter. In contrast, other studies have reported reductions in neutral detergent fiber following boiling or soaking treatments in D. giganteus, D. latiflorus, and D. sikkimensis (Kamthai, 2007; Rawat et al., 2016). The observed increase in our study may be attributed to the increased extractability of bound fiber fractions resulting from cell wall breakdown after heat treatment, hence enhancing the efficiency of enzymatic-gravimetric analysis. Conversely, studies indicating fiber reduction usually utilize extended boiling or other analytical techniques that may underreport fiber content due to the leaching of soluble fiber into the cooking water or the destruction of thermolabile fiber constituents. Furthermore, differences in bamboo species, maturation stage, and sample preparation methods (such as defatting and drying techniques) may lead to conflicting findings among research investigations.
Increased fiber intake is associated with improved glycemic regulation, as dietary fiber reduces glucose absorption and prevents postprandial blood sugar spikes, therefore reducing the glycemic index of foods (Anderson et al., 2009). Furthermore, dietary fiber is crucial for improving gut health by serving as a substrate for beneficial gut microbiota, which break down fiber to produce short-chain fatty acids (SCFAs) that enhance intestinal integrity and immune function (Slavin, 2013).
The total amino acid content followed the same trend as protein, with B. beecheyana containing significantly higher levels than D. latiflorus (1,609.50 mg/100 g vs. 924 mg/100 g, p<0.05; Table 2). Both species contained 18 amino acids, including all eight essential amino acids (EAAs). In B. beecheyana, aspartic acid was the most abundant (282±1.41 mg/100 g), followed by glutamic acid (255.5±9.19 mg/100 g), whereas tryptophan had the lowest concentration (20.5±0.71 mg/100 g). Similarly, D. latiflorus contained aspartic acid as the dominant amino acid (116.55±9.19 mg/100 g), closely followed by glutamic acid (116.4±4.24 mg/100 g), while methionine was the least abundant (10±0.00 mg/100 g).
After boiling, increases were observed in tyrosine, isoleucine, phenylalanine, leucine, lysine, and proline, ranging from 0.87% to 40.00%, with the greatest increase for methionine in D. latiflorus. In contrast, histidine, glycine, alanine, and arginine decreased consistently in both species (−2.42% to −30.00%), with histidine showing the largest reduction. The total amino acid content decreased in B. beecheyana (−1.46%) but increased in D. latiflorus (2.33%), as illustrated in Fig. 2.
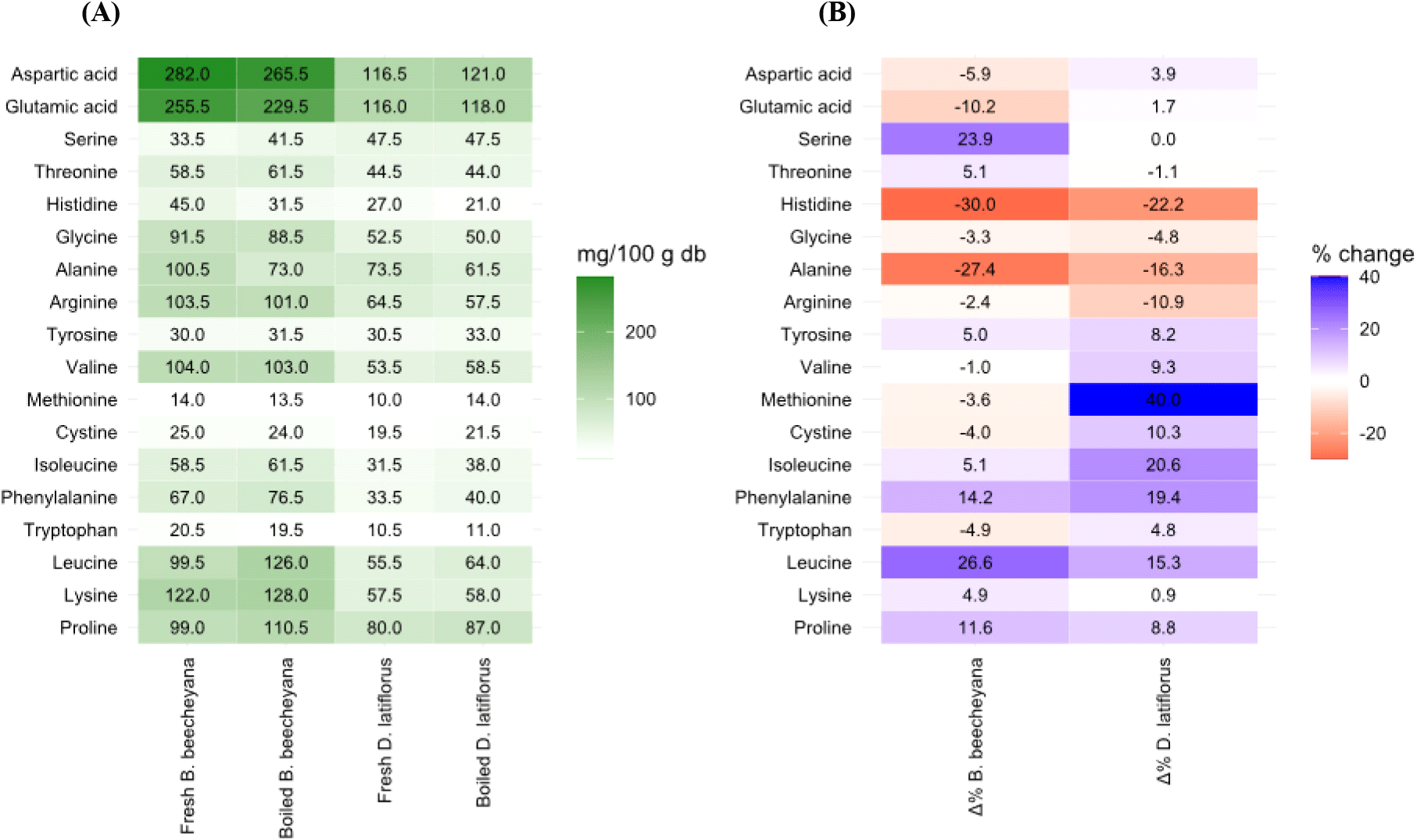
Similar reductions have been observed in other bamboo shoots, such as Phyllostachys praecox, following boiling or stir-frying, whereas steaming has been shown to preserve protein and amino acid content better (Zhang et al., 2011). This nutrient loss during boiling is not exclusive to bamboo; vegetables like cauliflower also exhibit significant protein losses when boiled. Comparative studies indicate that boiling causes more amino acid loss than roasting or microwaving, primarily due to the leaching of nutrients into the cooking water (Ito et al., 2019). Additionally, boiling induces protein denaturation, unfolding proteins and exposing hydrophobic regions, which promotes aggregation and potentially reduces amino acid bioavailability. Furthermore, heat exposure may chemically degrade labile amino acids, further contributing to overall protein reduction (Yu et al., 2015).
Protein quality was evaluated using the amino acid score (AAS) and the essential amino acid index (EAAI). The AAS compares the concentration of each EAA in the test protein to the FAO/WHO reference pattern, with values below 100% identifying limiting amino acids. The EAAI, by contrast, provides a holistic measure of protein quality by calculating the geometric mean of all EAA ratios relative to the reference (Machado et al., 2020). Significant varietal differences were observed, with B. beecheyana exhibiting a higher overall amino acid balance and superior protein quality compared with D. latiflorus (Table 3). Boiling further enhanced the AAS and EAAI values. This improvement may be attributed to the ability of thermal processing to disrupt cell wall structures and protein-matrix interactions, thereby increasing the extractability and detectability of amino acids. In addition, heat treatment may inactivate protease inhibitors or anti-nutritional factors, such as trypsin inhibitors, naturally present in bamboo shoots, thereby improving amino acid availability. Moist-heat treatments like boiling have also been shown to significantly reduce inhibitor activity in legumes and enhance protein digestibility (Ohanenye et al., 2022)
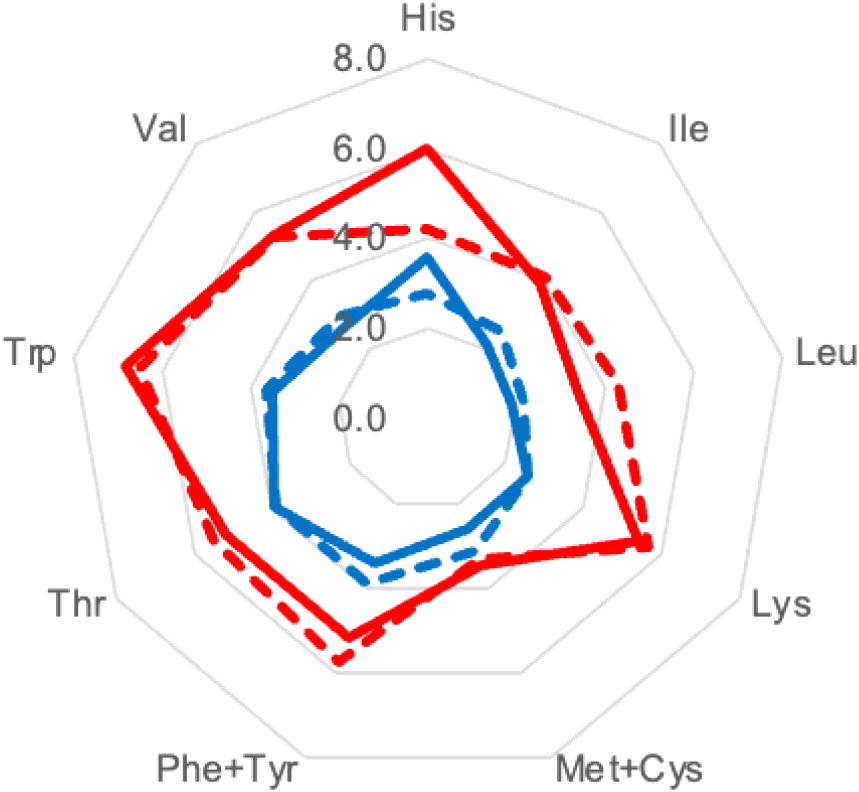
According to Thailand’s nutrition labeling regulations, the recommended daily intake of fresh vegetables is 80 g per day (Ministry of Public Health, 2023). Based on this consumption level, the essential amino acid contributions from bamboo shoots were compared with the WHO daily requirements for an average 60-kg adult. The highest contributions were observed for tryptophan (6.8% from fresh B. beecheyana), histidine (6.0% from fresh B. beecheyana), and lysine (5.4% from boiled B. beecheyana). In contrast, sulfur amino acids (cysteine + methionine, 2.6% in D. latiflorus and 3.5% in B. beecheyana) were identified as potential limiting amino acids in bamboo shoot protein (Fig. 3). Leucine and isoleucine were also identified as limiting amino acids, with contributions of 3.4-4.3% and 3.9-4.1% in B. beecheyana, and 1.9-2.2% and 2.1-2.5% in D. latiflorus, respectively. These findings indicate that, although bamboo shoots supply appreciable amounts of essential amino acids, their contribution at typical dietary intake levels is insufficient to serve as a sole protein source. This limitation can be addressed through dietary supplementation with cereals (e.g., rice, maize, wheat, quinoa) and seeds (e.g., sesame, sunflower), which provide relatively higher levels of sulfur amino acids, such as methionine, and also contribute branched-chain amino acids. Pairing bamboo shoots with legumes (e.g., soybeans, lentils, chickpeas) further enhances the supply of leucine and isoleucine (Singh et al., 2025).
Protein content was calculated using a nitrogen-to-protein conversion factor (NCF), which transforms nitrogen values obtained by the Kjeldahl or Dumas method into protein equivalents. The general factor of 6.25 has traditionally been applied to foods without specific conversion data, including bamboo shoots (Christian et al., 2015; Kusalaruk and Limsangouan, 2015). However, increasing evidence suggests that species-specific NCF values yield more accurate estimates of true protein content. For bamboo shoots, the average NCF calculated from the current study was 6.197 (Table 2), consistent with the range reported for other monocotyledonous plants (6.11-6.24) (Yeoh and Wee, 1994). Moreover, thermal processing may alter the NCF, as boiling can modify amino acid composition through leaching and heat-induced degradation, thereby influencing protein quality and the accuracy of protein estimations.
Phytosterols are plant-derived sterols structurally similar to cholesterol but with distinct physiological functions. Their most widely recognized clinical benefit is the reduction of low-density lipoprotein (LDL) cholesterol through competitive inhibition of intestinal cholesterol absorption. Daily consumption of 2-3 g of phytosterols has been shown to lower LDL cholesterol by approximately 10%, thereby contributing to a reduced risk of cardiovascular disease (Yang et al., 2019). In addition to their lipid-lowering effects, phytosterols have been linked to anti-inflammatory activity, antidiabetic properties, cancer-preventive potential, and immunomodulatory functions (Nattagh-Eshtivani et al., 2022).
In the present study, fresh B. beecheyana contained significantly higher phytosterol levels (4.66±0.18 mg/100 g) than fresh D. latiflorus (1.98±0.00 mg/100 g) (Table 4). Boiling resulted in an increase of approximately 44-46% in both species, reaching 6.73±0.14 mg/100 g in B. beecheyana and 2.89±0.08 mg/100 g in D. latiflorus. These values were below the interquartile range (IQR) previously reported for Bambusa and Dendrocalamus species (Fig. 1), suggesting that phytosterol content may vary according to species, geographic origin, cultivation conditions, and postharvest handling. Among individual phytosterols, β-sitosterol was predominant in both species, whereas campesterol and stigmasterol were detected in smaller amounts. Stigmastanol and brassicasterol were not detected in any of the samples.
The post-boiling increase in total phytosterols aligns with previous studies that reported enhanced phytosterol levels following the heat treatment of plant foods. For example, Kim et al. (2014) observed increased phytosterols in rice bran after microwave heating, while Moreau et al. (Moreau et al., 1999) reported similar effects in vacuum-oven-heated samples. Likewise, a study of nine vegetables, including bamboo shoots, documented increases in phytosterol levels (on a wet weight basis) after various cooking methods, with deep frying producing the largest increase (Shin et al., 2016). One possible explanation is that thermal processing disrupts plant cell walls and membrane structures, thereby improving phytosterol extractability from the plant matrix (Nartea et al., 2021).
Nonetheless, the relationship between heat treatment and phytosterol stability is complex. Some studies have reported that excessive heating, particularly at temperatures above 160°C, can induce oxidative degradation of phytosterols, resulting in the formation of phytosterol oxidation products that may diminish their health benefits (Kmiecik et al., 2020; Lin et al., 2018). These contrasting findings suggest that the impact of thermal processing on phytosterol content and bioavailability depends strongly on the heating method, temperature, and duration of treatment.
From a nutritional perspective, although bamboo shoots contain measurable amounts of phytosterols, the levels obtained from typical serving sizes (e.g., 80 g fresh weight/day, based on Thailand’s dietary reference intake for vegetables) remain far below the 2-3 g daily intake associated with clinically significant cholesterol-lowering effects. Therefore, bamboo shoot powders or extracts could serve as functional food ingredients that complement other sterol-rich matrices, such as cereal bran, nuts, or vegetable oils, to enhance total dietary phytosterol intake (Yang et al., 2019).
In food formulation, the bioaccessibility of phytosterols - i.e., their release and incorporation into mixed micelles during digestion - should be considered. Although the overall bioavailability of phytosterols in humans is low (0.5-2%), consuming approximately 3 g can still lower LDL-cholesterol by about 10.7% (Li et al., 2022). Bioaccessibility strongly depends on the surrounding food matrix. Specific components such as proteins and fats can markedly enhance sterol solubilization and absorption. For instance, complexing phytosterols with whey protein hydrolysate increased their in vivo absorption 2.84-fold compared to free phytosterols (Zhao et al., 2024). Embedding phytosterol nanoparticles in nanoporous starch aerogels increased bioaccessibility to 91.8% in regular-fat granola bars, compared to 16% in non-fat formulations (Ubeyitogullari and Ciftci, 2019). These findings indicate that, despite the relatively low phytosterol content in bamboo shoots, appropriate formulation strategies - such as combining bamboo shoot phytosterols with lipid- or protein-based carriers or using nanoscale delivery systems - could enhance their bioavailability and functional efficacy of phytosterols in bamboo shoots.
Phenolic compounds are important secondary metabolites in plants, contributing to protection against oxidative stress and offering health-promoting properties, including anti-inflammatory, antimicrobial, and anticancer effects. They also play a role in plant defense against biotic and abiotic stressors and act as natural antioxidants in the human diet (Ozcan et al., 2014). Bamboo shoots are recognized as a source of phenolic compounds, with levels influenced by species, maturity, and processing conditions (Joselin et al., 2014). In this study, fresh B. beecheyana exhibited significantly higher total phenolic content (TPC) and DPPH radical-scavenging activity than fresh D. latiflorus (Table 5). However, the TPC values of both species were below the interquartile range previously reported for Bambusa and Dendrocalamus species (Fig. 1). This discrepancy may reflect interspecific and geographical variability in phytochemical accumulation, differences in maturity stage, or postharvest handling.
Boiling significantly increased TPC in both species (91.4 to 144.7%) but slightly decreased DPPH radical-scavenging activity (−7.23 to −12.62%). Typically, phenolics are the primary contributors to hydrogen atom donation and free radical neutralization; therefore, a positive correlation between TPC and DPPH activity is often expected. However, in this study, TPC and DPPH values were negatively correlated (r= −0.669, p<0.05), suggesting that additional factors contribute to the overall reducing capacity. The Folin-Ciocalteu assay does not exclusively quantify phenolic compounds; rather, it reflects total reducing capacity, which can also include contributions from other reducing agents such as vitamin C, reducing sugars, and amino acids (Cheung et al., 2018). Therefore, the observed increase in TPC may result from the release of bound phenolics from the food matrix or the formation of new phenolic derivatives through structural rearrangements during boiling (Saikia et al., 2015). In contrast, the decline in DPPH activity indicates thermal degradation or structural modification of specific phenolic molecules. Flavonoids - particularly flavonols, flavones, and flavan-3-ols - are heat-labile and prone to oxidation, polymerization or depolymerization during cooking (Chaaban et al., 2017; Mohamed Ahmed et al., 2024).
Previous reports support the complexity of these interactions. For instance, boiling increased TPC in pepper but reduced it in squash, green beans, peas, and leek, with no significant changes in broccoli and spinach; antioxidant activity varied accordingly across species (Turkmen et al., 2005). Similarly, Pattarathitiwat et al. (2021) reported a reduction in the antioxidant activity of bamboo shoots after boiling. Together, these findings emphasize that thermal processing may enhance the apparent TPC of bamboo shoots by releasing bound phenolics and other reductants but simultaneously reduce radical-scavenging capacity due to degradation of key bioactive phenolic compounds.
Previous phytochemical analyses have identified several predominant phenolic compounds in bamboo shoots. Within the phenolic acid class, both hydroxybenzoic acids (e.g., p-hydroxybenzoic, protocatechuic, syringic, and vanillic acids) and hydroxycinnamic acids (e.g., chlorogenic, ferulic, and caffeic acids) have been reported. In the flavonoid class, the major representatives include the flavonol rutin and the flavan-3-ol catechin (Wang et al., 2020). These compounds exhibit broad antioxidant and anti-inflammatory activities, primarily through direct radical-scavenging mechanisms and by enhancing endogenous antioxidant enzyme systems. Chlorogenic, protocatechuic, and syringic acids have been linked to antidiabetic effects, while caffeic and vanillic acids have been exhibited cardioprotective properties. Additionally, caffeic, chlorogenic, and protocatechuic acids exhibit neuroprotective effects. Both hydroxybenzoic and hydroxycinnamic acids exhibit antimicrobial activity, which is attributed to their structural capacity to disrupt membranes. Flavonoids, particularly rutin and catechin, contribute further antioxidant, anti-inflammatory, and cardioprotective benefits (Oluwole et al., 2022).
Principal component analysis (PCA) was applied to visualize the variation in chemical composition among bamboo shoot samples. PCA is a multivariate technique that reduces data dimensionality while retaining most of the variation, enabling the identification of patterns and relationships among samples. In a biplot, the relative positioning of samples reflects similarities or differences in their composition, while the orientation and length of variable vectors indicate the degree of their contribution to each principal component (Wongsa et al., 2018). The PCA biplot (Fig. 3) revealed clear separation of bamboo shoot samples based on their chemical composition. The first two principal components explained 97.6% of the total variance, with PC1 (68.0%) strongly associated with moisture, total dietary fiber, fat, phenolics, and phytosterols, while ash and DPPH activity loaded negatively. This axis, therefore, reflects the compositional shifts induced by boiling, which increased moisture, fiber, fat, and phenolic contents but reduced ash and antioxidant activity. PC2 (29.6%) was dominated by protein (negative loading) and ash (positive loading), highlighting the trade-off between protein retention and mineral content across species and treatments. Fresh B. beecheyana was positioned closer to the protein and antioxidant vectors, consistent with its higher protein and radical-scavenging activity, while boiled samples shifted toward the moisture-fiber-lipid cluster. These findings demonstrate that PCA effectively captured nutritional and phytochemical changes associated with species differences and thermal processing.
4. Conclusions
This study provides the first comparative evaluation of the nutritional and bioactive profiles of B. beecheyana and D. latiflorus shoots in both fresh and boiled forms. B. beecheyana exhibited superior protein (41.68 g/100 g db), total amino acids (1,609.5 mg/100 g db), phytosterols (4.66 mg/100 g db), total dietary fiber (40.33 g/100 g db) and total phenolic content (204.42 mg GAE/100 g db) than D. latiflorus. Boiling significantly modified the chemical composition of both species, increasing moisture (1.4 to 2.0%), dietary fiber (16.8 to 22.6%), fat (128.9 to 228.8%), phytosterols (44.4 to 45.9%), and total phenolics (91.4 to 144.7%), while decreasing protein (−0.3 to −62.3%), ash (−40.6%), and antioxidant activity (−7.2 to −12.6%). Overall, boiled B. beecheyana provided the most favorable nutritional profile while minimizing potential health risks associated with cyanogenic compounds and other natural toxins present in raw bamboo shoots.
Although both species supplied notable amounts of essential amino acids―particularly tryptophan, histidine, and lysine―methionine, cysteine, leucine, and isoleucine remained in limited supply. To enhance their nutritional value, combining bamboo shoots with cereals or seeds rich in sulfur- and branched-chain amino acids could improve amino acid balance and overall protein quality. The predominance of β-sitosterol also suggests potential cardioprotective benefits.
As fast-growing, underutilized species, bamboo shoots represent a sustainable and nutrient-dense food source suitable for integration into functional foods and plant-based diets. The findings of this study offer novel insights into species-specific nutrient profiles, thermal stability, and functional potential, thereby supporting future research on optimizing processing conditions and enhancing bioavailability for health-oriented and sustainable food systems.
