
1. Introduction
Exposure to cadmium (Cd), a toxic heavy metal, occurs through inhalation, ingestion, and dermal contact, with occupational exposure being a common concern. Cd exhibits toxicity across multiple targets, such as the liver and kidney (Genchi et al., 2020; Matović et al., 2015). Earlier in vivo explorations have substantiated the intricate correlation between Cd-triggered cytotoxicity and apoptosis in the hepatic tissues of both murine and rat models (Habeebu et al., 1998; Tzirogiannis et al., 2003). Moreover, parallel in vitro examinations have further affirmed that Cd elicits the fragmentation of DNA—an unmistakable hallmark of apoptosis—within HepG2 cells, a lineage of human hepatocellular carcinoma cells (Aydin et al., 2003; Fatur et al., 2002; Shimoda et al., 2001).
The involvement of key molecular players, such as p53, Bcl-2 family proteins, caspases, and MAPK signaling pathways, has been identified in Cd-induced apoptosis. Cd activates p53, a tumor suppressor protein, which promotes apoptosis through the regulation of pro-apoptotic genes. The balance between anti-apoptotic (e.g., Bcl-2) and pro-apoptotic (e.g., Bax) members of the Bcl-2 family influences apoptotic cell fate. Caspases, as central executioners of apoptosis, cleave downstream targets to execute cell death. Furthermore, MAPK signaling pathways, including JNK, p38, and ERK, have been implicated in Cd-induced apoptosis.
In recent years, in vitro cell-based models have played a vital role in toxicological research, enabling the assessment of Cd toxicity and elucidating underlying mechanisms. Numerous studies utilizing HepG2 cells have shed light on the mechanisms underlying Cd-induced hepatotoxicity, including oxidative stress, mitochondrial dysfunction, apoptosis, and inflammation. These cellular responses contribute to liver damage and may lead to the development of liver diseases.
Antioxidants and antioxidant enzymes such as N-acetylcysteine, glutathione, catalase, vitamin E, and superoxide dismutase are effectively applied to reduce Cd-induced hepatotoxicity (Aronis et al., 2003; Mattie and Freedman, 2001; Poliandri et al., 2003; Pourahmad and O’Brien, 2000). Crocus sativus and green tea extract had also been reported to inhibit Cd-induced hepatocyte toxicity in adult rats (Abdelrazek et al., 2016; Yari et al., 2016). Additionally, research has explored the detoxifying effects of Rhizoma coptidis (RC) aqueous extract and the intracellular distribution and metabolism of ginsenosides in animal livers (Park et al., 2013).
Elaeagnus umbellata, also known as autumn olive, is a deciduous shrub cultivated in South Korea. The fruit and leaves of this plant are edible. The fruit, known as “Mokbanha” in traditional Korean medicine, has been reported to have effects on conditions such as diarrhea (Rafique et al., 2016), bleeding, and indigestion (Lee et al., 2011). It is also known to contain approximately 7-8 times more lycopene than tomatoes (Wang et al., 2020). Studies have reported various pharmacological activities of E. umbellata leaves, including anti-bacterial, anti-inflammation (Kang et al., 2020), skin whitening and wrinkle-improving effects (Park et al., 2019), and anticancer activity (Kim et al., 2016). However, no studies have reported the efficacy of the extract from E. umbellata leaves against hepatotoxicity caused by Cd.
Although research on the major active components of E. umbellata leaves is still limited, it is known to contain gallic acid, kaempferol, catechin, and other compounds (Abri and Maleki, 2016; Yoon et al., 2007). Gallic acid is widely recognized for its antioxidant properties (Yen et al., 2002) and has been reported to exhibit anti-inflammatory activity (Kroes et al., 1992), anti-allergic effects (Kim et al., 2006), inhibit lung cancer proliferation (Ohno et al., 1999), and inhibit the growth of oral bacteria (Kang et al., 2008). The exceptional antioxidant properties of green tea, which is rich in gallic acid, are well known. Similarly, kaempferol, a phenolic component abundant in plants, has been investigated for its anti-inflammatory activity (Devi et al., 2015) and its ability to alleviate oxidative stress in nerve cells (Sharma et al., 2007).
In this study, we aimed to investigate the cellular protective effects of ethanol extract from E. umbellata leaves (EUL) against Cd-induced cytotoxicity in HepG2 cells. Moreover, the potential of the extract to prevent reactive oxygen species (ROS) levels, apoptosis, and Cd accumulation induced by Cd treatment was also examined.
2. Materials and methods
Cadmium chloride [CdCl2], 4′,6-Diamidine-2′-phenylindole dihydrochloride (DAPI), 2′,7′-dichlorofluorescin diacetate (DCF-DA), 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were purchased from the Sigma-Aldrich Co. (St. Louis, MO, USA). Antibodies against phospho-extracellular signal regulated kinase 1/2 (ERK; #9101), phospho-protein kinase B (Akt; #9271), phospho-c-Jun N terminal kinase 1/2 (JNK; #9251), phospho-p38 (#9211), β-Actin (#3700), Caspase-3 (#14220), Phospho-p53 (#9284), Nuclear factor erythroid-derived 2-related factor 2 (NRF2; #12721), Heme oxygenase-1 (HO-1; #26416), superoxide dismutase type 1 (SOD-1; #2770), Catalase (#12980), Metallothionein (MT)-1,2, anti-mouse secondary antibody (#7076), and horseradish peroxidase linked anti-rabbit secondary antibody (#7074) were purchased from Cell Signaling Technology (Beverly, MA, USA). Dulbecco’s modified Eagle’s medium (DMEM) and fetal bovine serum (FBS) were from the Welgene (Gyeongsan-si, Korea). All other chemicals and reagents used were of analytical grade.
The preparation of EUL was adapted from the method previously described by Kang et al. (2020). E. umbellata plants cultivated in Hapcheon-gun, Gyeongsangnam-do, Korea, were acquired for the study. The dried leaves of E. umbellata were subjected to extraction using 70% ethanol (Duksan Science, Seoul, Korea) at a ratio of 10 times its weight. The extraction process was carried out at a temperature of 80°C for a duration of 6 h. Subsequently, the obtained extract was concentrated and then subjected to lyophilization using a freeze-dryer (FDU-2100, EYELA, Tokyo, Japan), resulting in the production of EUL. The lyophilized EUL was confirmed to yield approximately 16.8%.
Human hepatoma HepG2 cells were cultured in DMEM, which was supplemented with 10% heat-inactivated FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin. The cells were maintained in a controlled environment, specifically at a temperature of 37°C in a 5% CO2-95% air incubator with regulated humidity levels. In all experimental setups, a control group was incorporated, functioning as the vehicle control group. This control group was treated with 0.1% DMSO.
The assessment of cell viability was carried out using the MTT assay method. HepG2 cells were seeded in a 96-well plate using DMEM and were cultured at 37°C for a duration of 24 h. Following the incubation period, the cells were subjected to treatment with varying concentrations of EUL ranging from 0 to 500 μg/mL. Additionally, cells were exposed to 10 μM CdCl2 concurrently with escalating concentrations of EUL (0-500 μg/mL). Upon the completion of the specified incubation period, a solution of 20 μL of MTT (1 mg/mL) was introduced to the cells. This solution was allowed to incubate with the cells for a duration of 4 h. Subsequent to the incubation, the culture media were aspirated and replaced with 100 μL of DMSO. The mixture was gently agitated for 10 min to facilitate complete dissolution. The absorbance of the resultant solution was measured at a wavelength of 540 nm utilizing a microplate reader (Model: CA 94089, Molecular Devices, San Jose, CA, USA).
HepG2 cells were seeded in a 6-well plate at a density of 1×106 cells per well, utilizing DMEM as the culture medium. The cells were then cultured at a temperature of 37°C for a duration of 24 h. Following the incubation period, HepG2 cells were subjected to a 24 h treatment with CdCl2 and EUL. Post-treatment, HepG2 cells were lysed utilizing lysis buffer (Pro-Preptm, iNtRON, Seongnam, Korea) supplemented with a protease inhibitor (P3100_001, GenDEPOT, Barker, TX, USA) and a phosphatase inhibitor (P3200_001, GenDEPOT). The lysed cells were subsequently incubated on ice for 30 min. Protein concentrations within the cell lysates were determined employing a BCA protein assay (Thermo Fisher Scientific, Waltham, MA, USA). Following protein quantification, 20 μg of proteins were loaded and subjected to separation through 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The separated proteins were subsequently transferred onto PVDF membranes (Whatman GmbH, Dassel, Germany). The membranes were blocked with a solution of 5% skim milk in Tris-buffered saline containing 0.1% Tween-20 for a period of 1 h. Subsequent to blocking, the membranes were incubated with primary antibodies for 16 h at 4°C. Following primary antibody incubation, the membranes underwent three successive washes using Tris-buffered saline containing 0.1% Tween-20. Subsequently, horseradish peroxidase-linked secondary antibodies (Cell Signaling Technology) were incubated with the membranes for 1 h. Protein visualization was achieved via enhanced chemiluminescence, and the images were captured and analyzed using UVP Vision Works® LS Image Acquisition & Analysis Software (Upland, CA, USA).
For the visualization of nuclear morphological alterations and reactive oxygen species (ROS) in Cd-treated cells, a double staining procedure utilizing DAPI and DCFDA dye was employed. The treated cells were fixed using a solution of 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS) for a duration of 10 min. Subsequently, the cells were subjected to three consecutive PBS washes and were then incubated with a combination of 0.1 mg/mL DAPI and 25 μM DCF-DA for 10 min at room temperature. Following the staining process, the cells were washed thrice with PBS. The levels of ROS within the cells were visualized and analyzed using a fluorescence microscope. The quantification of intra-cellular CdCl2 levels was directly performed via ICP-MS and is expressed as parts per billion (ppb) per 1×106 cells.
HepG2 cells cultivated in a complete culture medium were employed to quantify the levels of Cd, and these cells were designated as the Cd-control group. Post the cultivation period, cells were subjected to two washes with phosphate-buffered saline (PBS) to eliminate any potential metal contamination originating from the culture medium. The washed cells were subsequently centrifuged (95 ×g, 10 min), resuspended in PBS, and a specified volume of each sample was enumerated using a hemocytometer to achieve a consistent count of 1×106 cells. The collected cells were then suspended in 180 μL of royal water (composed of nitric acid at a ratio of 1:3 with hydrochloric acid) and subsequently lysed for a duration of 30 min. The washing solutions along with the royal water utilized for cell dilution underwent analysis through Inductively Coupled Plasma Mass Spectrometry (ICP-MS) using an Agilent 7500cx instrument (Agilent, CA, USA). Calibration procedures were executed utilizing Cd standard solutions (Sigma-Aldrich Co.).
The results are expressed as means±standard deviation ellipse (SDE) values, derived from three independent experimental replicates. To determine the significance of differences between each experimental test condition and its respective control, Student’s t-test was employed for each paired experiment. Significance levels were designated at *p<0.05, **p<0.01, and ***p<0.001. Statistical analyses were performed by using Microsoft Excel software (Microsoft Co., Redmond, WA, USA).
3. Results and discussion
We first examined the cell viability of HepG2 cells incubated with EUL (50-500 μg/mL) in both the presence or absence of Cd to determine the protective effect of the extract against Cd toxicity (Fig. 1(A) and (B)). No cytotoxicity was observed by EUL treatment, but the cell viability was decreased to 34.3% after 24 h Cd (10 μM) exposure. Pretreatment with EUL show an increase of cell viability dose-dependently, indicating hepatoprotective effect of the extract.
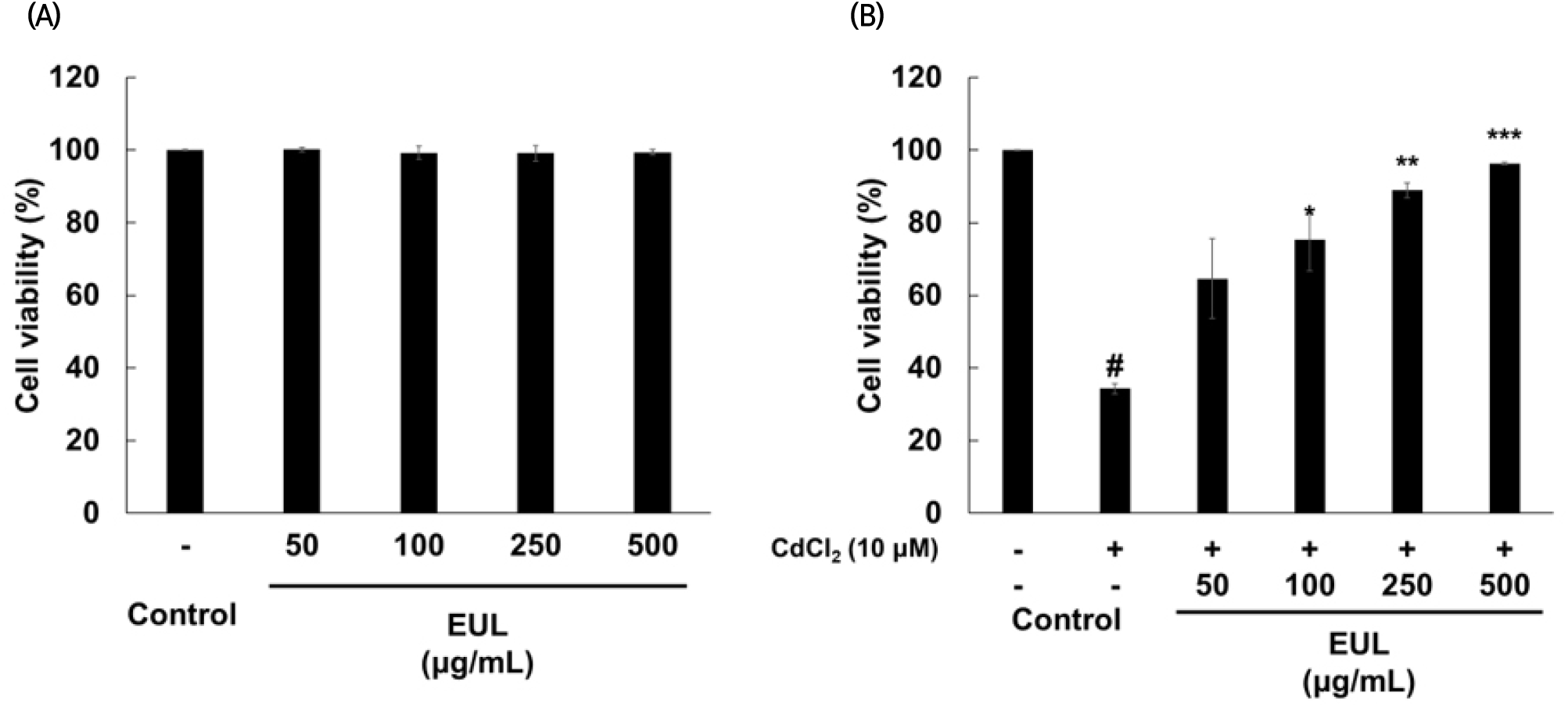
In this study, the concentration of Cd applied to HepG2 cells was deliberately chosen to achieve a cytotoxic effect exceeding 50%, thereby enabling the evaluation of the preventive efficacy of EUL at different concentrations. Consequently, a concentration of 10 μM was employed, resulting in approximately 63% cytotoxicity. Furthermore, pretreatment with EUL before the introduction of Cd exhibited a significant inhibitory effect on Cd-induced cytotoxicity in HepG2 cells. This protective impact against Cd-induced cytotoxicity in our study seems to be attributed to the prominent presence of antioxidant compounds in EUL, notably gallic acid, as evidenced in various investigations on the hepatoprotective properties of polyphenolic compounds in plant extracts against oxidative stress on HepG2 cells (Martin et al., 2008; Zhang et al., 2020). Previously, our research had identified that gallic acid and kaempferol as the primary constituents in the leaf extract of E. umbellata, containing 98.95 mg/100 g and 81.85 mg/100 g, respectively (Kim et al., 2016).
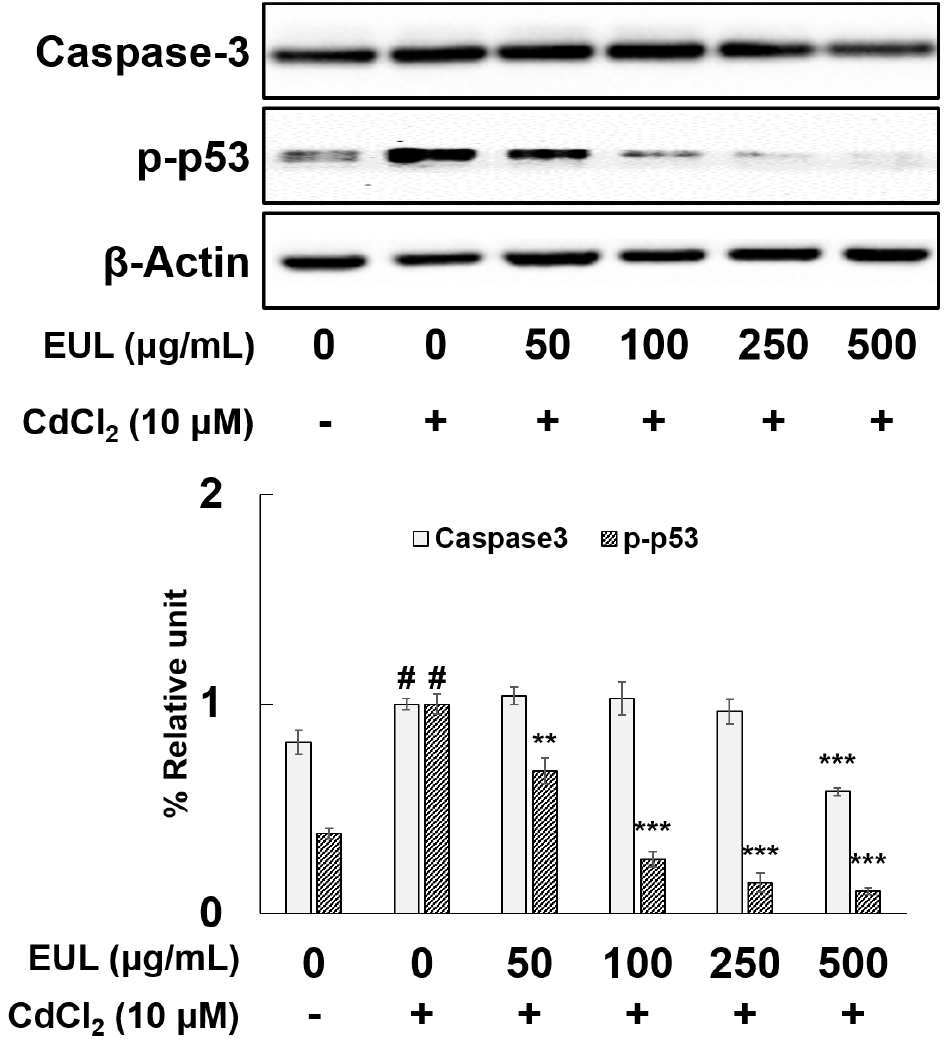
In light of the pivotal role played by caspase activation and phosphorylated p53 (p-p53) in the initiation of apoptosis in response to diverse stimuli, our investigation aimed to elucidate the alterations in the levels of caspase-3 and p-p53 within HepG2 cells upon exposure to Cd. Additionally, we sought to examine the potential protective effects of EUL against Cd-induced apoptotic cell death. To achieve this, the cells underwent pre-treatment with various concentrations of EUL (50, 100, 250, 500 μg/mL) for 24 h, followed by subsequent exposure to Cd for an additional 24 h. Our assessment of protein expression unveiled a significant increment of approximately 1.2-fold in caspase-3 levels and a more pronounced elevation of approximately 2.6-fold in p-p53 upon Cd exposure (as depicted in Fig. 2). Remarkably, pre-treatment with EUL at the concentration of 500 μg/mL exhibited a substantial mitigating effect. This resulted in a reduction of 42% in the expression of caspase-3 and a noteworthy decrease of 74% in p-p53 levels compared to Cd-treated control cells. These outcomes collectively validate the engagement of apoptosis-associated proteins in the course of Cd-induced cell demise within HepG2 cells. Furthermore, the observed protective influence of EUL, particularly in attenuating the Cd-induced alterations in these proteins—especially p-p53—affirms its role in safeguarding against the protein changes triggered by Cd. Our study corroborates earlier findings that have implicated Cd in the initiation of apoptotic cell death within HepG2 cells (Athmouni et al., 2018; Lawal et al., 2015; Oh and Lim, 2006). Notably, the use of Z-DEVD-FMK, a potent caspase 3 inhibitor, effectively thwarted Cd-induced cell death, thereby highlighting the role of caspase 3 in cell demise at these levels. Caspases are categorized into two distinct groups: ‘apoptotic initiators’, represented by caspase 8, and ‘apoptotic effectors’, exemplified by caspase 3 (Oh and Lim, 2006). Caspase-3 activation can be initiated through the death receptor pathway, culminating in apoptotic cell death (Alarifi et al., 2013). Furthermore, exposure of HepG2 cells to Cd has been reported to trigger the phosphorylation and upregulation of the p53 protein (Seo et al., 2013). This, in turn, orchestrates the transcription of downstream proteins, including p21 and Bax, pivotal players in cellular processes linked to apoptosis (Oh et al., 2006).
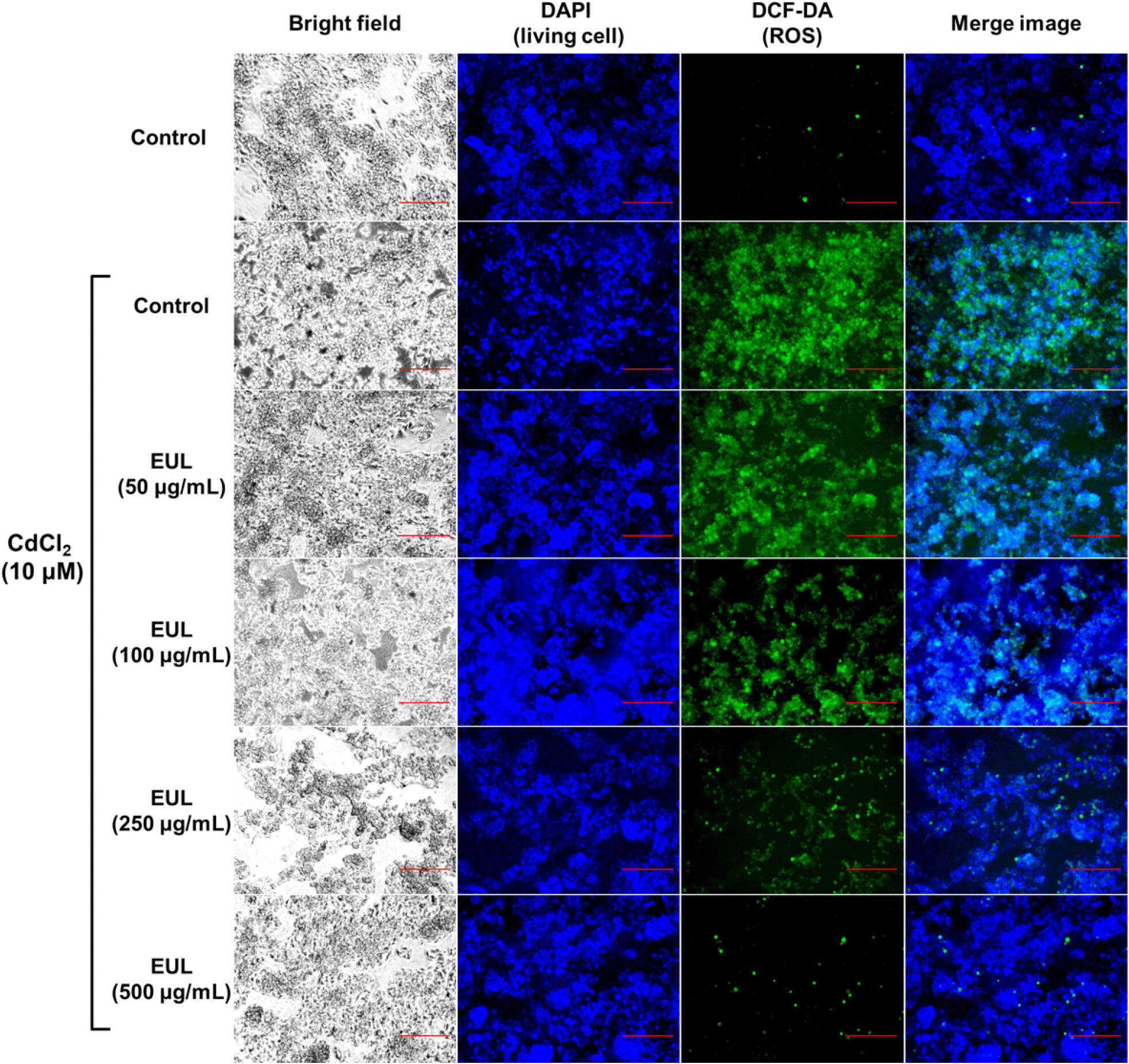
Given the well-established literature on Cd-triggered ROS production in hepatocytes (Fotakis et al., 2005), our study sought to qualitatively explore the regulatory impact of EUL on ROS levels within Cd-treated HepG2 cells. Utilizing microscopy and H2DCF-DA staining, we aimed to ascertain whether fluctuations in intracellular ROS levels correlated with the apoptosis instigated by Cd exposure. Specifically, the effect of Cd treatments (10 μM for 24 h) was assessed, revealing a notable surge in DCF fluorescence an indicator of increased ROS accumulation within HepG2 cells in comparison to untreated control cells (Fig. 3). However, cells that were pre-treated with EUL exhibited a relatively diminished DCF fluorescence signal when contrasted with Cd-treated cells. Notably, the pre-treatment with EUL, specifically at concentrations of 250 and 500 μg/mL, displayed significant effectiveness in curbing ROS levels relative to cells solely subjected to Cd exposure. Notably, the control cells displayed no discernible DCF fluorescence. These findings underscore the potential hepatoprotective attributes of EUL, positing its role in mitigating ROS generation incited by Cd. Although ROS were traditionally associated with detrimental biological effects due to their propensity to induce oxidative harm to cellular constituents, it has been subsequently recognized that ROS assume a pivotal role in normal biological processes. This encompasses participation in signaling pathways governing diverse transcription factors and subsequent gene expression (Hsiao and Stapleton, 2004). It is pertinent to note that several reports have substantiated the effectiveness of free radical scavengers in impeding apoptosis initiated by Cd, acting through their antioxidative attributes. In essence, Cd triggers an initial ROS surge, consequently instigating the activation of the redox-sensitive MAPK pathway (Papaiahgari et al., 2004).
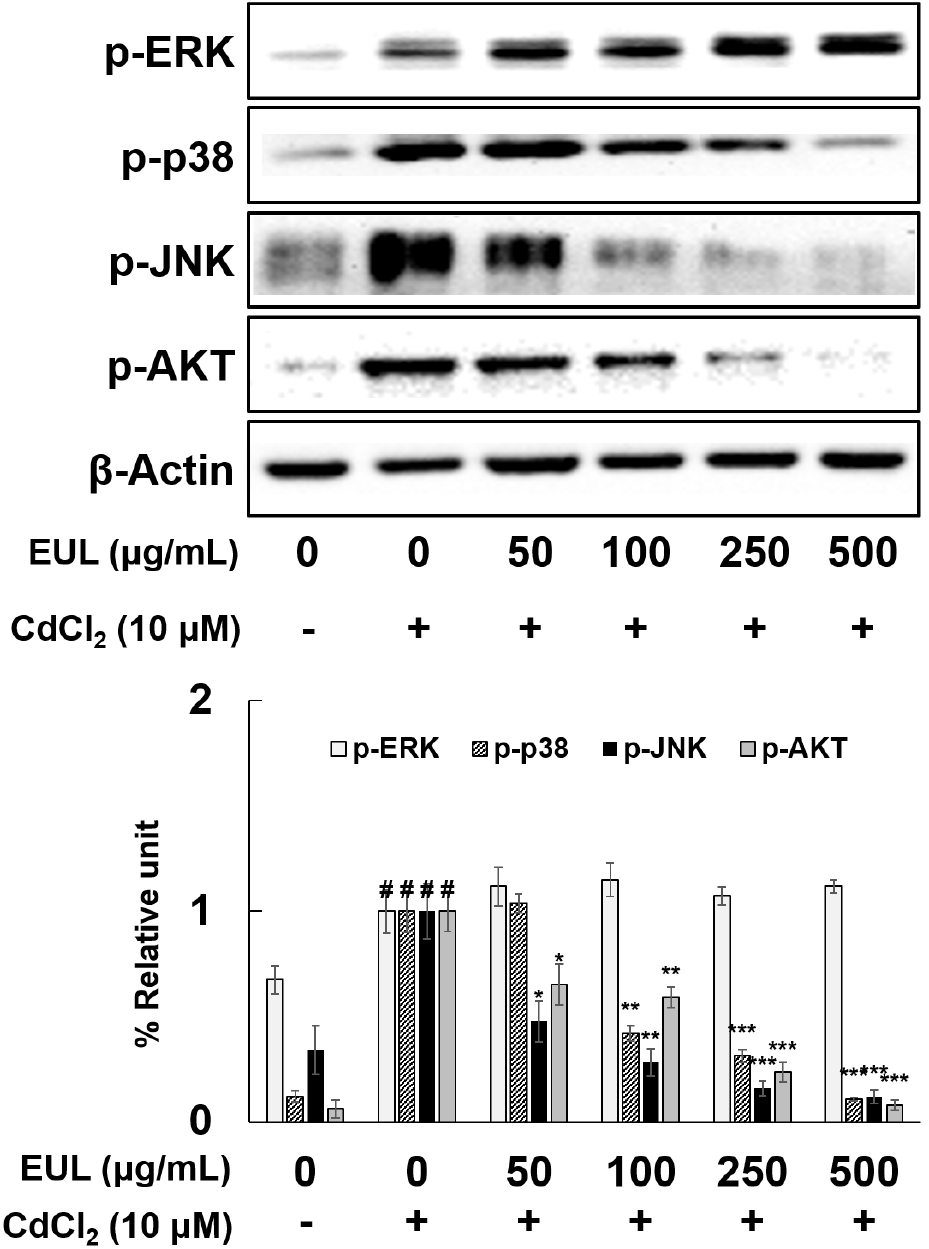
Cd activation of various signal transduction factors, including Akt, ERK, p38, and JNK in diverse cell types, including HepG2 cells, has been well-documented (Escobar et al., 2009; Tsai et al., 2016). Furthermore, Shen et al. (2004) highlighted the role of MAPK pathways in triggering the expression of antioxidant genes through a Nrf2-dependent mechanism. In order to ascertain the contribution of MAPKs to the protective effect exerted by EUL against Cd-induced cytotoxicity in HepG2 cells, we examined the activation states of AKT, JNK, ERK, and p38 within the cells. Upon treatment of HepG2 cells with Cd (10 μM) for a duration of 24 h, phosphorylation levels of ERK, p38, JNK, and AKT exhibited significant increments of approximately 1.5-fold, 8.3-fold, 2.9-fold, and 16.7-fold, respectively (as depicted in Fig. 4). Notably, pretreatment with EUL (50-500 μg/mL) for a period of 24 h markedly suppressed the Cd-induced escalation in phosphorylation of p38, JNK, and AKT in a dose-dependent manner. Intriguingly, the phosphorylation status of ERK remained unaffected following EUL pretreatment. These outcomes collectively reaffirmed that the heightened phosphorylation of MAPKs is a hallmark of Cd-induced apoptosis, and further substantiated that EUL selectively counteracted the phosphorylation of JNK, p38, and AKT. Interestingly, the phosphorylation of ERK remained unaltered. This study thus establishes the pivotal role of Cd-induced MAPK phosphorylation in mediating apoptosis, with phosphorylated MAPKs transmitting apoptotic signals to the nucleus through the activation of sub-signaling molecules, as previously noted (Tsai et al., 2016). The current investigation reinforces the finding that Cd-triggered phosphorylation of MAPKs is modulated by EUL, specifically inhibiting the phosphorylation of JNK, p38, and AKT. Consequently, EUL emerges as a regulator that suppresses Cd-induced apoptosis within HepG2 cells. Future research endeavors involving MAPK inhibitors are warranted for a comprehensive understanding of this intricate mechanism.
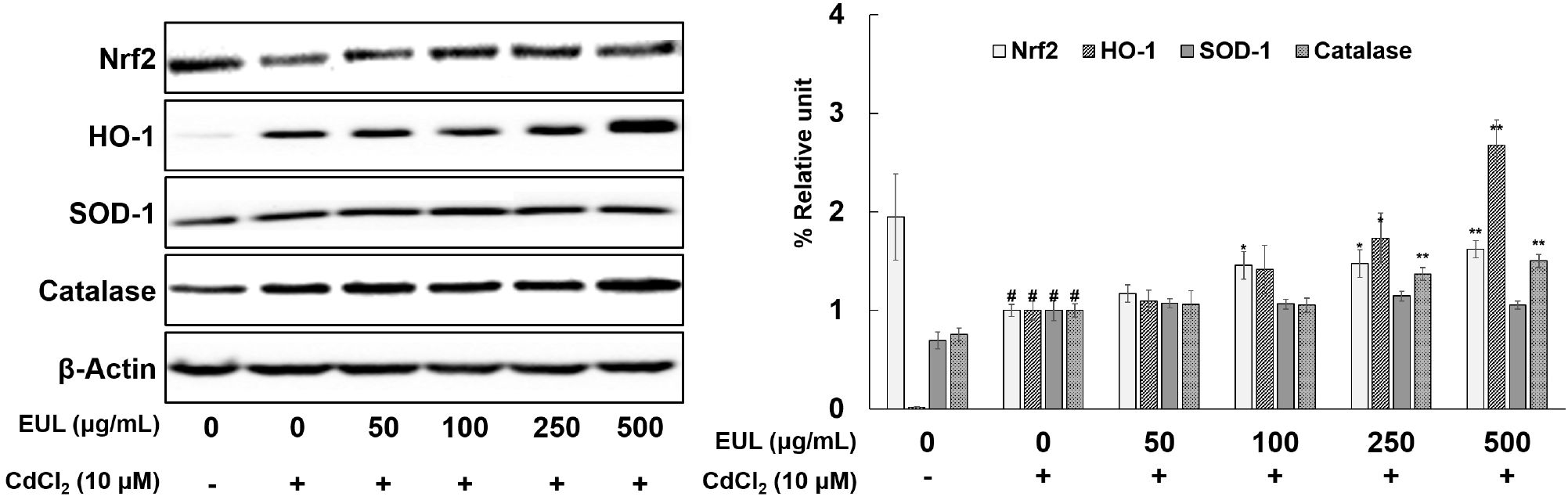
To comprehensively explore the effect of EUL on the levels of Nrf2 and its downstream target molecules, namely HO-1, SOD-1, and catalase, in the context of Cd-induced toxicity within HepG2 cells, we conducted a rigorous analysis of protein expressions through Western blotting. As demonstrated in Fig. 5, relative to the untreated-control cells, exposure to Cd resulted in an approximate 50% reduction in Nrf2 protein expression, concomitantly with a substantial elevation in HO-1 expression. Furthermore, Cd exposure induced notable increases in SOD-1 (1.4-fold) and catalase (1.3-fold) expression levels (Fig. 5). Pre-treatment with EUL (500 μg/mL) led to a significant elevation in the protein levels of Nrf2 (1.6-fold), HO-1 (2.7-fold), and catalase (1.5-fold) in comparison to the Cd-exposed control cells. SOD-1 expression, however, remained unchanged across all tested concentrations. Our findings underscore the dose-dependent augmentation of Nrf2 expression mediated by EUL, thereby mitigating the suppression induced by Cd exposure. Additionally, the protein expression of both HO-1 and catalase demonstrated an up-regulated pattern following treatment with 10 μM Cd for 24 h. Remarkably, these elevations were further enhanced by EUL pre-treatment in HepG2 cells, underscoring the potential of EUL to mitigate Cd-induced hepatotoxicity by up-regulating key antioxidant proteins. Given the established role of Nrf2 as a pivotal cytoprotective transcription factor orchestrating the transcription of target genes encoding an array of antioxidant enzymes, our results align with the existing understanding that various antioxidant compounds exert their hepatoprotective effects by activating the Nrf2 signaling pathway (He et al., 2008; Jadeja et al., 2016; Lee et al., 2012).
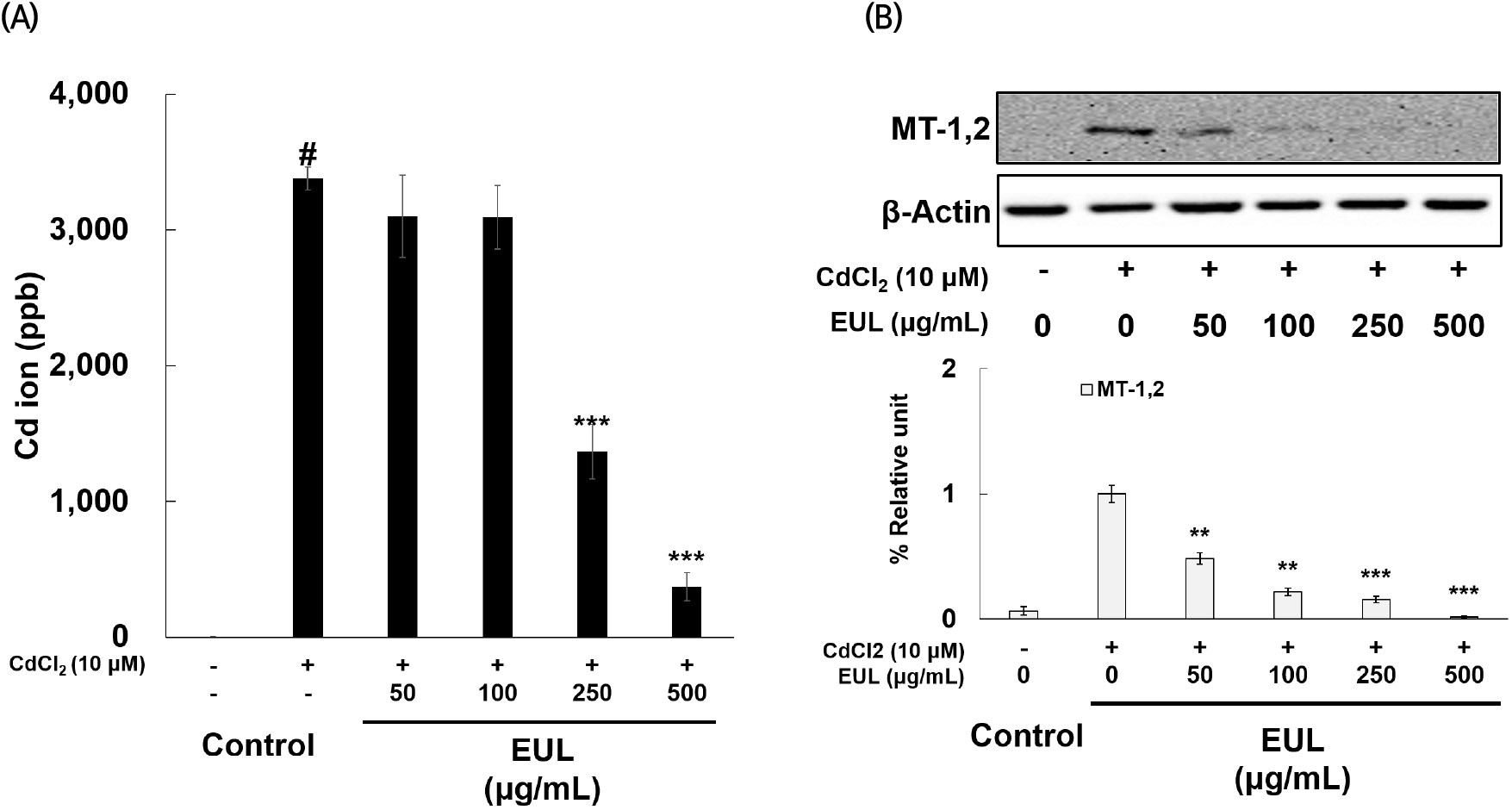
Intracellular Cd levels were investigated via rigorous ICP-MS analysis in cells exposed to Cd. The quantification of Cd levels within these cells serves as a proxy for Cd absorption or uptake by the cultured cells. Consequently, we assessed intracellular Cd levels following exposure to 10 μM Cd in HepG2 cells, which revealed an observable elevation in intracellular Cd levels. After 24 h of Cd exposure, 3,379 ppb of Cd were detected in one million HepG2 cells (as shown in Fig. 6(A)). This finding unequivocally underscores the ability of HepG2 cells to effectively uptake Cd, affirming its capacity to traverse the cellular membrane. Intriguingly, pre-treatment with EUL at concentrations of 250 and 500 μg/mL significantly attenuated the heightened Cd concentration within the cells, reducing it by approximately 60% and 11%, respectively, compared to Cd-exposed control cells. Furthermore, the expression of metallothionein (MT)-1,2 exhibited a marked upregulation upon exposure to 10 μM Cd, relative to untreated control cells. Remarkably, EUL pre-treatment induced a progressive decline in MT expression (as depicted in Fig. 6(B)). This observation corroborates existing knowledge that Cd stimulation leads to the induction of genes encoding MTs. Notably, MTs play a pivotal role in orchestrating protective mechanisms during Cd exposure of target organs, as previously documented (Urani et al., 2007). Collectively, our data suggest that EUL effectively mitigates Cd-induced oxidative stress by curbing the cellular uptake of Cd. This critical action ultimately contributes to hepatoprotection against Cd-induced cellular damage. Although HepG2 cells are widely used as a hepatocyte model, they may not be completely identical to normal hepatocytes. Therefore, comparative experiments with normal hepatocytes or other model systems may be necessary to ensure the accuracy of the results.
4. Conclusions
In the present study, we explored the potential protective effects of EUL against toxicity induced by Cd in HepG2 cells. Our observations revealed an augmentation in cell death, an upregulation of apoptotic protein expression, and an accumulation of ROS in response to Cd treatment, indicative of heightened oxidative stress. Notably, pretreatment of the cells with EUL resulted in an enhancement of antioxidant protein expression, in line with the notably robust antioxidant activity previously reported for this extract. In summary, our current data strongly imply that EUL effectively counteracts Cd-induced hepatotoxicity by impeding the activation of MAPK pathways (p38, JNK, AKT), promoting Nrf2 activation and the consequent upregulation of antioxidant proteins such as HO-1 and catalase. Additionally, EUL appears to downregulate MT expression by mitigating ROS-induced oxidative stress. Therefore, these findings underscore the potential utility of EUL as a food material, offering protective attributes against hepatic damage attributed to Cd exposure in humans.
