서 론
염증반응은 다양한 세포에서 분비되는 산화적 스트레스 물질이나 감염성 물질에 의하여 유발되는 정상적인 방어기전으로써 한시적으로 일어나고 빠르게 회복된다(Kim 등, 2000). 그렇지만 다양한 염증성 질환들에서 염증반응과 면역이 적절하게 감소되지 않고, 염증성 면역세포에 분비되는 ROS, 과도한 염증성 cytokine 및 chemokine이 대장 및 지방을 포함하여서 여러 조직에 만성적인 염증반응을 일으키는 것으로 알려져 있다(Mases, 2008). Chemokine 중에서도 monocyte chemoattractant protein 1(MCP-1)은 CC chemokine으로써 단핵구, 림프구, 호염구 등을 선택적으로 유도하며, interleukin-1β (IL-1β), tumor necrosis factor-α(TNF-α), 저밀도 지단백질 (LDL) 등에 반응하여 기질세포, 사구체의 내피, 세뇨관 상피세포, 모세혈관의 내피세포 및 평활근세포 등에서 생산된다고 보고되고 있다(Kim 등, 2002). Nuclear factor-kappa B (NF-κB)는 염증 반응, 세포 증식, 면역 및 세포 사멸과 관련된 유전자의 신호전달 및 발현에 관여하는 유도성 전사인자이며(Park과 Hong, 2016), NF-κB 경로는 TNF receptor superfamily members(TNFRs), T-cell receptor(TCR), B-cell receptor(BCR)와 TNF-α, interleukin-1β(IL-1β)와 같은 염증성 cytokine에 의해 유발된다(Barroso 등, 2016). NF-κB family transcription factor complexes는 p50, p52, p65(ReIA), ReIB 및 c-Rel과 같은 NF-κB 단위 구성원에 의해 형성된 동종 또는 이종이량체로 구성되어 있으며, 핵 내로 이동하여 TNF-α, IL-6, IL-8, IL-1β 및 MCP-1와 같은 염증 반응에 관여하는 다기능 cytokine의 전사를 조절한다. 또한 arachidonic acid를 prostaglandin E2 (PGE2)로 전환시켜 피부 염증을 유발하는 염증 효소인 cyclooxygenase-2(COX-2)의 발현과 염증 매개체인 NO와 NO를 합성하는 inducible nitric oxide synthase(iNOS)의 발현을 유도한다. iNOS 과발현은 만성 염증, 인간 종양의 발생 및 발암과도 관련이 있다(Kuo 등, 2015; Lindstrom과 Bennett, 2005). 그 중에 nitric oxide(NO)는 여러 염증 반응에 핵심적인 매개 물질이다(Lee 등, 2008). NO는 혈류와 혈압을 조절, 혈소판의 응집을 억제, 신경전달과정 등에 관여하여 인체의 방어기전과 면역조절로서 중요한 역할을 하고, 염증상태에서 과도하게 NO가 생성되면 부종, 혈관투과성 등 염증반응이 촉진되며, 염증 매개체의 생합성이 촉진되어 급성 또는 만성염증에 대한 반응이 나타나 조직 손상 및 유전자 변이 등이 나타날 수 있다(Ryu 등, 2003). PGE2의 합성은 phospholipase A2의 효소작용에 의해 막 인지질(membrane phospholipid)로부터 arachidonic acid가 만들어지는 것으로 시작된다. Arachidonic acid는 효소작용에 의해 PGG2가 되고, 다시 불안정한 대사산물인 PGH2가 되는데, 이 두 과정은 COX에 의해 촉진된다(Ji 등, 2004). Eicosanoinds의 생합성 과정 중에서 COX에 의해 염증 반응이 진행되며, 유효성 효소들과 염증성은 COX-2에 의해 나타난다(Gordon, 1998). 염증반응에 중요 역할을 하는 PGE2는 COX-2에 의하여 형성되며, 염증 반응의 매개체로서의 역할뿐만 아니라(Bishop 등, 2002), Th2 type 면역반응을 촉진하고 Th1 type의 면역반응은 억제하며, 대식세포에서 TNF-α, IL-1β, IL-8, IL-12 등의 pro-inflammatory의 생성을 억제하고, 항염증성 cytokine의 생성을 촉진하는 면역반응의 조절자로서의 역할이 최근 수년간의 연구에서 밝혀진 바 있다(Harris 등, 2002).
석위(Pyrrosia lingua)는 상록성 양치식물로서 고란초과 (Polypodiaceae)에 속한다. 우리나라 남부지방, 제주도 등 따뜻한 지역의 산지 바위나 나무껍질에 분포하여 자생하는 여러해살이풀이다(Do 등, 1992; Yamashita 등, 1998). 한방에서는 지상부의 전초를 말려 통증, 요로결석 형성 및 질 출혈에 유용한 효과가 있는 한약재로 사용되고 있다. 이 식물에 대한 이전의 식물 화학적 연구에서 chlorogenic acid, xanthonoids, kaempferol, isoorientin, organic acids, xanthones, triterpenoids, steroids 및 volatile oil 등이 조사되어 알려져 있으며(Chen 등, 2011), 항바이러스, 정균, 이뇨제, 면역강화 및 신장보호 활성 등의 많은 기능성 효과를 가지고 있다고 보고된 바 있다 (Kim 등, 2010; Lim 등, 2014). 그러나 이러한 석위가 약리적 효능을 보유하고 있을 가능성에도 불구하고, 현재까지는 석위의 항산화 및 진통 효능 등만 보고되고 있으며(Kim 등, 2017), 아직까지 석위의 염증 억제에 관한 과학적 연구가 이뤄지지 않았고, 그를 밝히기 위한 수행 연구가 필요한 실정이다.
본 연구에서는 Raw 264.7 cell에서 NF-κB 신호 전달 조절을 통해 cytokine의 발현량을 조절하기 위해 Pyrrosia lingua ethanol extract(PLE)를 처리하여 NF-κB p-65, iNOS, COX-2 단백질을 확인한 후, 이들의 pro-inflammatory mediators인 prostaglandin E synthase 2(PTGES2)와 nitrite의 발현량, cytokine 발현 수준을 통해 염증 억제 연구를 수행하였다.
재료 및 방법
실험에 사용한 세포는 mouse 유래의 macrophage인 Raw 264.7 cell(Korean Cell Line Research Foundation)을 구매하여 사용하였다. 사용한 시약은 Dulbecco’s modified Eagle’s medium(DMEM)(HyClone Laboratories, Inc., Logan, Utah, USA), inactivated fetal bovine serum(FBS)(HyClone Laboratories, Inc.), penicillin-streptomycin(HyClone Laboratories, Inc.), dimethyl sulfoxide(DMSO)(Junsei Chemical Co., Tokyo, Japan), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)(Sigma-Aldrich Co., Louis, MO, USA), phosphate buffer saline(PBS)(HyClone Laboratories, Inc.), mammalian protein extraction reagent(M-PER)(Thermo Fisher Scientific, Waltham, MA, USA), nuclear and cytoplasmic extraction reagents(NE-PER)(Thermo Fisher Scientific.), protease inhibitor (Thermo Fisher Scientific.), BCA kit(Thermo Fisher Scientific.), polyvinylidene fluoride(PVDF) membrane(Millipore Corp, Bedford, MA, USA) 등의 시약을 구입하여 사용하였다.
Western blot을 측정하기 위한 시약은 iNOS(1:1,000 in 5% skim milk; #sc-7271; Santa Cruz Biotechnology, Dallas, TX, USA), COX-2(1:500 in 5% skim milk; #33345; Signalway antibody, Maryland, College Park, USA), NF-ĸB p65(1:1,000 in 5% skim milk; #6956T; Cell Signaling Technology, Danvers, MA, USA), p-NF-ĸB p65(1:500 in 5% skim milk; #3036S; Cell Signaling Technology), glyceraldehyde 3-phosphate dehydrogenase(GAPDH)(1:1,000 in 5% skim milk; #MA5-15738; Thermo Fisher Scientific.), β-actin(1:500 in 5% skim milk or 5% BSA; #sc-47778; Santa Cruz Biotechnology) antibody와 mouse anti-rabbit lgG-HRP(1:1,000; sc-2357; Santa Cruz Biotechnology), goat anti-mouse IgG(1:1,000; 31430; Thermo Fisher Scientific.) secondary antibody를 구입하여 사용하였다.
Real-time polymerase chain reaction(PCR)을 측정하기 위한 시약은 GeneAll®Ribospin RNA extraction kit(GeneAll Biotechnology Co, Seoul, Korea), qPCRBIO cDNA synthesis kit(PCR Biosystems, London, UK), Real time PCR master mix 5 μL(GeneAll Biotechnology Co.)를 구입하여 사용하였다.
본 실험에 사용된 기기는 동결건조기(freeze dryer, FD8518, Ilshinbiobase, Yangju, Korea), rotary vacuum evaporator (Eyela NE, Tokyo, Japan), CO2 incubator(311, Thermo Fisher Scientific.), heating block(MaXtable H10-Set, Daihan scientific co., Ltd., Wonju, Korea), enzyme-linked immunosorbent assay(ELISA) reader(SPECTRO star Nano, BMG LABTECH, Ortenberg, Germany), transfer cell 기기(Hofer, Holliston, MA, USA), Azure Biosystems(C300, Azure Biosystems inc., Dublin, CA, USA), PCRmax Eco 48 real-time PCR system (PCRmax, Staffordshire, UK) 등을 사용하였다.
본 연구에서 사용된 Pyrrosia lingua는 서울약령시장 ㈜천지가약초(Seoul, Korea)에서 2017년에 구입하여 사용하였다. 재료로 사용된 Pyrrosia lingua 잎은 이물질을 제거한 후, 45°C dry oven(Jeiotech, Daegu, Korea)에서 건조한 후, 고속분쇄기(RT-08, Rong Tsong Precision Technology, Taichung, Taiwan)를 이용하여 25,000 rpm으로 분쇄하여 40 mesh 채에 걸러 powder 형태로 제조한 다음 4°C에서 저온저장하며 사용하였다. 실험에 사용한 추출물은 Pyrrosia lingua 잎 분말에 80% ethanol 용액(1:100)을 첨가하여 24시간 동안 shaking incubator에서 실온으로 교반 추출하였다. 추출물은 filter paper(No.1, Whatman Inc., Piscataway, New Jersey, USA)로 여과한 후 rotary vacuum evaporator를 이용하여 농축하여 유기용매를 제거하였다. 농축된 추출물은 동결건조 한 후 DMSO에 용해하여 −20°C시료 보관 냉장고에서 저온 보관하며 실험에 사용하였다.
Mouse 유래의 macrophage인 Raw 264.7 cell을 10% FBS와 1% penicillin-streptomycin을 첨가한 DMEM 배지를 이용하여 37°C의 5% CO2 incubator를 이용하여 배양하였다.
Pyrrosia lingua ethanol extract(PLE)가 세포에 독성을 일으키는지 평가하기 위해 MTT assay로 확인하였다. Raw 264.7 cell(5×103 cells/mL)을 각각 48 well plate에 분주하여 24시간 전 배양한 후, PLE를 5-100 μg/mL 농도별로 첨가한 배양액을 처리하고 18시간 배양하였다. 이후 well plate에 5 mg/mL MTT 시약 50 μL를 첨가하여 4시간 동안 반응시키고 배양액을 제거한 후, 남아있는 formazan을 DMSO로 용해시켜 ELISA reader를 이용해 540 nm에서 흡광도를 측정하였다. 시료의 독성이 정상상태의 Raw 264.7 cell에 영향을 미치는지 확인하기 위해 세포의 수와 형태학적 변화를 현미경 (×400, ECLIPSE TS100, Nikon, Tokyo, Japan)으로 관찰하였다.
Lipopolysaccharide(LPS)로 자극된 세포로부터 생성된 nitrite에 대한 PLE가 미치는 영향을 알아보기 위해 griess reagent system’s kit 시약(1% sulfanilamide + 0.1% naphthylendiamine dihydrochloride, 1:1) 반응을 이용하여 배양액 중에 존재하는 NO2-의 형태로 측정하였다. Raw 264.7 cell(5×104 cells/mL)을 96 well plate에 분주하고, 37°C의5% CO2 incubator에서 24시간 전 배양하였다. 세포에 1 μg/mL의 LPS와 PLE를 농도별로 처리한 배양액으로 18시간 재배양하였다. Nitrite 생성량은 supernatant를 모아 griess reagent system’s kit를 사용하여 반응시킨 후 ELISA reader를 이용하여 540 nm에서 흡광도를 측정하였다. 세포 배양액 내 nitrite의 농도는 0.1 M sodium nitrite(NaNO2)의 농도별 표준곡선에 대입하여 산출하였다.
LPS로 자극된 Raw 264.7 cell(5×105 cells/mL)의 COX-2, iNOS protein 발현량 측정을 위해 6 well plate에 세포를 분주하고, 24시간 전 배양하여 세포에 1 μg/mL LPS로 자극을 준 다음, PLE를 농도별(20-50 μg/mL)로 처리한 배양액으로 24시간 재배양하고 PBS로 3회 세척하였다. M-PER와 protease inhibitor를 섞은 mixture를 각 well당 70 μL 첨가하여 4°C에서 lysis시킨 후, 16,000 ×g에서 20분간 저온 원심 분리기(4°C, Gyrozen, Seoul, Korea)를 이용하여 세포막 성분 등을 제거하였다. 원심 분리하여 얻은 supernatant는 BCA kit로 정량하였으며, 총 20-30 μg의 단백질을 10%의 sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE)를 통해 100 V로 1시간 30분간 전기 영동하여 분리하였다. 분리된 단백질은 transfer cell 기기를 이용하여 60 V로 2시간 30분간 PVDF membrane으로 transfer한 후 실온에서 1시간 동안 blocking하였다. Blocking이 끝난 membrane에 COX-2, iNOS, GAPDH antibody를 사용하여 4°C에서 over night한 후, buffer를 이용하여 3회 세척하였다. Goat anti-mouse IgG secondary antibody를 1시간 동안 반응시킨 후 1×TBST buffer로 3회 세척하였으며, enhanced chemiluminescence (ECL) kit(Millipore Corp.)와 반응시켜 Azure Biosystems를 이용하여 band를 현상하고, Image J software(NIH-Scion, USA)를 사용하여 정량하였다.
LPS로 자극된 Raw 264.7 cell(2×106 cells/mL)이 되도록 분주하고, 5% CO2 incubator에서 37°C로 24시간 동안 배양하였다. 교체된 배지에 LPS(1 μg/mL)로 자극을 준 다음 PLE를 농도별로 첨가한 배양액을 처리하고 30-60분 동안 배양하였다. 배양 후, 세포를 수집하고 차가운 PBS로 3회 세척하였다. Cytoplasmic과 nuclear 단백질 추출은 제조업체의 프로토콜에 따라 NE-PER를 사용하여 준비하였다.
LPS로 자극된 Raw 264.7 cell(5×106 cells/mL)의 TNF-α, IL-1β, IL-6, MCP-1, prostaglandin E synthase 2(PTGES2) mRNA 발현량 측정은 100×20 mm culture dish에 세포를 분주하고 24시간 전 배양하여 1 μg/mL의 LPS를 처리하고 PLE를 농도별로 처리한 배양액으로 24시간 재배양하고 PBS로 3회 세척하였다. mRNA 발현량을 비교 분석하기 위해 제조업체의 프로토콜에 따라 total RNA를 GeneAll®Ribospin RNA extraction kit를 사용하여 추출하였으며, qPCRBIO cDNA synthesis Kit를 이용하여 cDNA를 합성하였다. 합성된 cDNA는 PCRmax Eco 48 real-time PCR system을 이용하여 유전자별로 mRNA 발현량을 실시간 분석하였으며, PCR condition과 primer sequence는 Table 1, 2와 같다.
결과 및 고찰
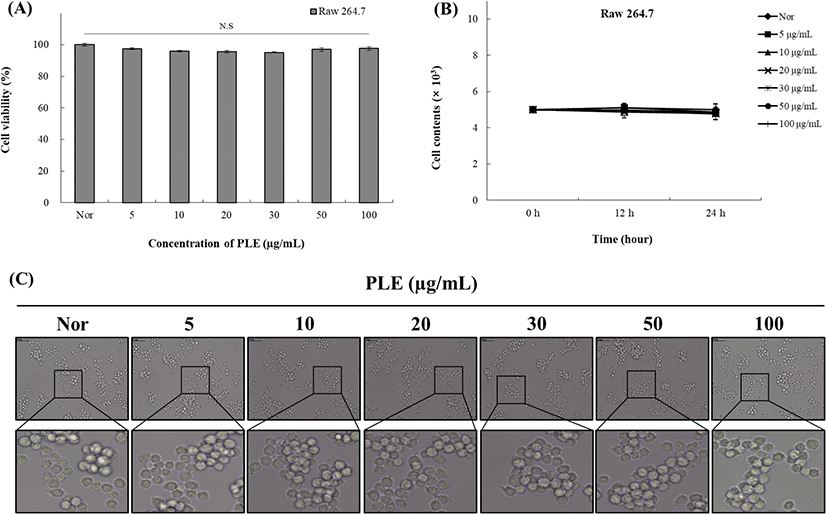
PLE의 Raw 264.7 cell에 대한 독성을 확인하기 위해 MTT assay를 이용하여 세포 생존율을 측정한 결과는 Fig. 1(A)와 같다. 정상상태의 Raw 264.7 cell에 PLE를 5, 10, 20, 30, 50, 100 μg/mL의 농도별로 처리하였을 때 PLE 처리군에서는 농도가 증가하여도 세포 생존율이 유의적으로 감소하는 경향이 나타나지 않았으며, 100 μg/mL의 고농도로 처리하였을 때도 세포 생존율이 97.7%로 확인되어 고농도의 PLE 처리 시에도 시료가 세포에 독성을 나타내지 않는 것으로 확인되었다. 이러한 결과를 확인하기 위해 시료를 처리한 후 0, 12, 24시간 후에 현미경을 통해서 세포의 수를 세고(Fig. 1(B)), 형태학적 변화도 관찰하였다(Fig. 1(C)). 그 결과는 Fig. 1(A)에서와 같이 PLE 처리군에서 MTT assay에서 나타난 세포독성의 경향과 유사한 결과를 나타내었다(p<0.01). 위의 결과에 따라 PLE는 Raw 264.7 cell에 대한 세포독성을 가지지 않음을 확인하여 100 μg/mL를 최고 농도로 설정하여 추후 실험에 적용하였다.
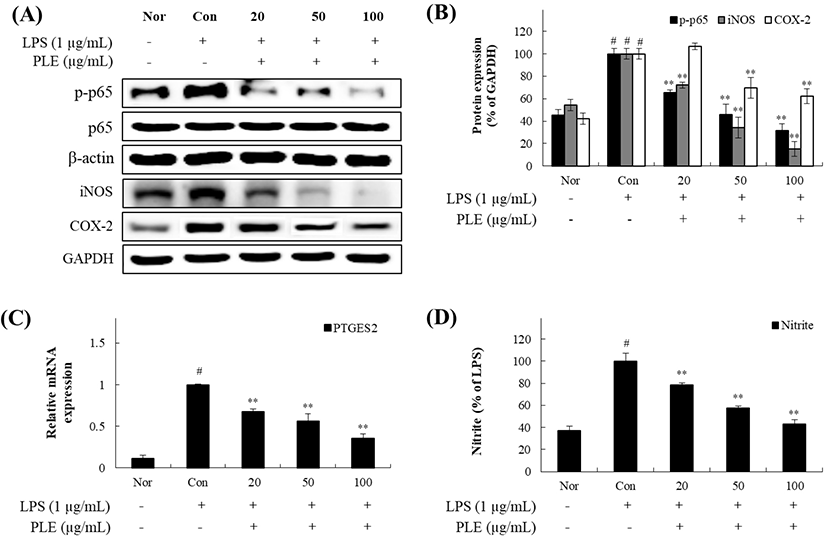
면역반응의 일종인 염증은 외부 자극, 병원체 또는 손상된 조직의 침입에 대한 방어적 보호 메커니즘이다(Oh 등, 2013). 외부 자극에 의한 염증은 주로 NF-κB 신호전달경로에 의해 조절된다. NF-κB는 면역체계 조절(immune modulation), 상피세포의 분화(epithelial differentiation) 및 염증반응 조절 등에 관여하는 단백질군(protein family)으로 유전자의 발현을 조절하여 세포 내 신호전달 체계를 이루는 인자이다(Shin 등, 2012). 비활성 상태의 NF-κB 복합체는 세포질에서 IκB-α에 결합되어 통제되지만, LPS 유도로 인해 활성화되어 proteasome을 통한 ubiquitination을 매개하고, IκB-α의 분해를 유도하여 조절한다(Cheung 등, 2013). 이어서, NF-κB 복합체는 세포질에서 핵으로 이동하여 iNOS, COX-2 유도성 효소단백질의 생성 및 염증성 cytokines 생성에 관여하는 유전자의 promoter에 결합하여 암호화하며, 이들의 생성을 촉진한다(Rehman 등, 2012). 따라서 Raw 264.7 cell에서 염증성 cytokines 발현에 직접적으로 관여하는 NF-κB 신호 경로를 조사하기 위하여 LPS 유도된 Raw 264.7 cell에 PLE(20, 50, 100 μg/mL)를 농도별로 처리하여 NF-κB complex 중 인산화 된 p65 protein과 NF-κB 활성화에 의해 생산된 염증매개물질인 iNOS, COX-2의 발현량을 확인하였다. 그 결과, Fig. 2(A)와 2(B)에서와 같이 농도별 PLE 처리군에서 cytoplasmic 대비 nuclear 내 p-p65 발현량이 유의하게 감소하였으며, 염증매개물질인 iNOS, COX-2 발현량 측정 결과에서도 p-p65 단백질 발현량 억제 양상과 유사하게 농도에 따라 유의하게 조절하였다. 한편, NOS와 COX-2는 주로 염증의 초기 반응에 관여하는데, 이들에 의한 pro-inflammatory mediators인 PTGES2와 nitrite의 발현량을 측정한 결과, Fig. 2(C)와 2(D)에서와 같이 COX-2에 의해 매개되어 PGE2 생성에 관여하는 PTGES2의 mRNA 발현량과 iNOS mediators인 nitrite 발현량이 PLE 처리에 의해 유의적으로 감소되는 것으로 조사되었다. 이상의 결과에 따라 PLE는 PTGES2와 nitrite를 조절하여 이들의 전사인자인 iNOS와 COX-2에 대한 직접적인 억제력과 LPS로 유도된 NF-κB 신호전달경로의 단계적인 반응을 조절한다고 판단되었다. 따라서 염증반응에 직접적인 영향을 미치는 인자인 염증성 cytokine의 발현에도 관여할 것으로 판단되어 이에 대한 반응을 조사해 보였다.
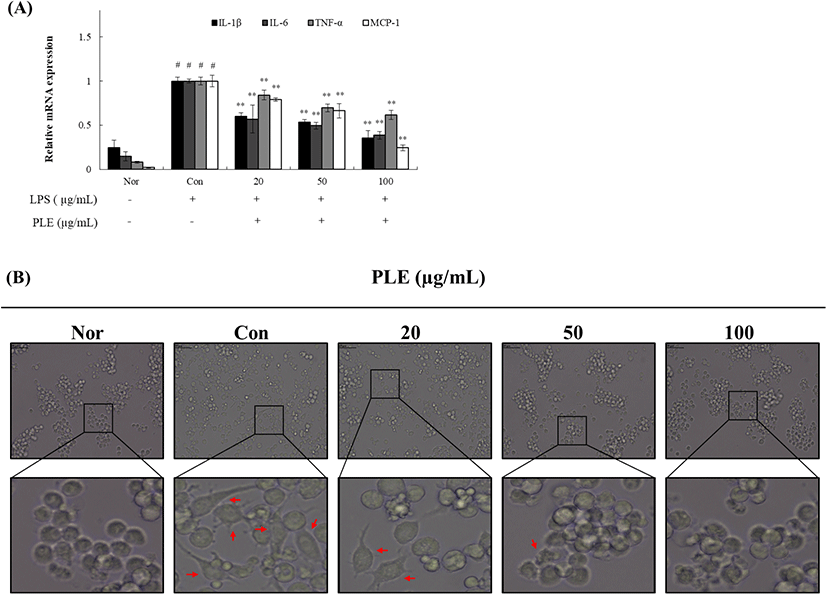
NF-κB는 monocytes/macrophages에서 pro-inflammatory cytokines의 유도를 매개한다. 이러한 cytokine의 대부분은 선천성 면역세포 및 섬유아세포에서 NF-κB를 활성화하여 추가적인 염증성 cytokine 및 chemokine의 발현을 유도하여 염증성 면역세포의 증식과 급성 염증 및 만성 염증의 전염을 유도한다(Vannier, 1992). 염증에 의한 cytokine에는 TNF-α, IL-6, IL-1β 등이 존재하며, 탐식세포를 포함한 면역세포에서 만들어진다. 이들은 모두 국소적으로 작용하여 응고를 유도하고 혈관투과성을 증가시키며, COX-2, iNOS의 생성을 유도하는 일들을 수행한다(Bazarsad, 2012). 따라서 NF-κB에 의하여 조절되는 cytokines과 chemokine인 IL-1β, IL-6, TNF-α, MCP-1에 대해 조사하기 위하여 LPS 유도된 Raw 264.7 cell에 PLE(20, 50, 100 μg/mL)를 농도별로 처리하여 IL-1β, IL-6, TNF-α, MCP-1 발현량을 확인하였다. 그 결과, Fig. 3(A)와 같이 농도별 PLE 처리군에서 L-1β, IL-6, TNF-α, MCP-1의 mRNA 발현량이 처리 농도가 증가할수록 유의하게 감소하였다. LPS를 처리하지 않은 Normal(Nor)군과 세포에 LPS만 처리한 Control(Con)군, LPS 처리 후 농도별 PLE 처리군의 세포의 형태적인 변화를 관찰한 결과는 Fig. 3(B)와 같다. 처리 후 Nor군에서는 부착성 세포인 macrophage의 형태적 특징인 구형의 모양인 반면, Con군에서는 LPS를 처리함에 따라 노화된 세포의 불규칙적인 모양과 손상된 구조의 뾰족뾰족하게 뻗은 형태를 나타내었다. LPS 처리 후 농도별 PLE 처리군의 경우, 세포의 형태적인 변화가 Nor군의 상태보다는 손상된 구조와 노화된 세포가 조금 관찰되었지만, 농도별 PLE의 처리에 따라 세포의 부착성과 손상된 구조가 Con군에 비해 다소 감소하는 것을 확인하여 이는 PLE의 처리 농도에 따라 형태적 차이를 보인다고 판단하였다. 종합하면, PLE는 외부의 자극에 의해 손상을 받는 macrophage에 대해 NF-κB 신호 경로와 cytokine의 발현량을 감소시키는 것을 확인하였으며, 따라서 PLE가 염증반응에 관여하는 인자들을 하향 조절하는 것으로 판단할 수 있었다. 따라서 본 연구에서는 PLE가 염증반응을 저해하는 기능성 식품 소재 및 항염증 치료제로서의 가능성과 가치를 확인하였으며, 염증 억제에 대한 잠재력 뿐만 아니라, 국내에 자생하는 우수 생물자원에 대한 적극적인 발굴 연구 및 유용 생물자원의 활용성 증대를 위한 기술 개발 산업에서 유익한 응용 가능성을 제시하는 연구임을 시사하는 바이다.