








서 론
당장법(sugaring)을 적용한 생물재료의 삼출액이 식품 등 여러 분야에 이용되고 있다. 당장법은 당의 삼투압을 이용하여 생물 조직 내부의 물과 유용성분의 추출 및 미생물 생육을 억제함과 동시에 감미를 부여하는 저장 방법으로 그 과정에 발생하는 액상의 부분을 삼출액이라고 한다. 이것은 당절임 삼출액(Park 등, 2020), 당침출액(Park 등, 2020), 식물발효액 (Han 등, 2012), 발효 추출액(Kim 등, 2011), 청(Kim 등, 2020), 효소식품(Kim 등, 2015; Lee와 Lee, 2001) 등 다양한 이름으로 불리고 있다. 이 당장법에 의한 삼출액은 현재 식품공전 중 농수산 가공식품류의 효소식품 항목에 해당할 수 있으나, 당장법과는 관계없이 일정량 이상의 조단백질과 효소를 함유하는 등 규격 기준에 적합해야 하며, 그 외에도 제품의 종류에 따라 발효 음료류 등에 포함될 수 있다.
제조하는 방법은 일반적으로 과일이나 채소 등의 생물성 식품 재료에 동량의 당을 혼합하여 실온에서 저장하는 것이다(Kim 등, 2013). 이 혼합물은 저장 기간이 증가함에 따라 생물조직이 파괴되어 액즙의 형성 등 외관의 변화, 가스의 생성, 질감이나 냄새, 그리고 색깔 변화 등 여러 면에서 대폭 변하는데 이 과정을 발효로 표현하기도 한다. 따라서 이러한 삼출액의 저장은 단지 식품의 가식 기간을 연장하는 의미의 저장 외에 지속적으로 식품이 제조되는 과정을 포함한다. 한편, 삼출액의 제조에는 보통 설탕이 사용되나, 올리고당(Kim 등, 2011)이나 꿀(Kim 등, 2020)이 사용되기도 한다.
이러한 당장법에 따른 삼출액은 원료에 따라 그 기능성이 다양하다. 대표적인 것이 다시마, 양파, 알로에, 매실, 케일 등이다. 이들 원료 중 다시마는 항산화 효과, 항돌연변이, 항균 활성 등의 생물학적 활성을 갖고 있다(Okai 등, 1993; Park 등, 2009; Wang 등, 2006; Yoo 등, 2002). 또한, 양파는 지질에 대한 항산화 효과(Bracco 등, 1981; Kwak 등, 2000), 항암작용(Jeong 등, 2009), 항돌연변이 활성(Gwon 등, 1999), 혈중 콜레스테롤 감소(Kim과 Chun, 1999), 고혈압 및 당뇨병에 대한 효과(El-Demerdash 등, 2005; Yang 등, 2008) 등 중요한 생리활성을 가지는 것으로 알려져 있다. 알로에는 항염증 효과를 나타내며(Cho 등, 2006), 매실은 항산화 활성 (Hwang 등, 2004; Park 등, 2002; Xia 등, 2010), 항균력(Ha 등, 2006; Ko와 Yang, 2009), 항암 효과(Bae 등, 2000; Chuda 등, 1999) 등이 있는 것으로 보고되었다. 케일은 발암 억제 효과(Cheigh 등, 1998; Kim 등, 1999; Park, 등, 1992)와 항암성(Hertog 등, 1992)이 알려져 있다.
그러나 시판 당장법에 의한 이들 삼출액에 대하여는 명확한 정의와 규격, 제조 기술의 과학화, 고품질 상품화, 식품의 안전성 확보 등의 면에서 충족되지 못한 부분이 있어 고시형 건강기능 식품 유형에서 제외되었다(Han 등, 2012). 따라서 본 연구에서는 당장법에 의한 삼출액의 과학적인 이해를 위해 다시마, 양파, 알로에, 매실 및 케일을 원료로 하여 설탕을 이용한 당장법에 의한 삼출액을 만들고, 1년 동안 저장하는 동안 주요 화학적 성분의 변화를 확인하고, 우점종 젖산 생성 세균을 분리 및 동정하고, 이에 대한 미생물학적 특성을 조사하였다. 또한, 전통적인 항아리 저장 방법을 대신하여 스텐리스강 용기가 사용될 수 있는지도 알고자 하였다.
재료 및 방법
삼출액의 제조에는 ㈜건강을지키는사람들(나주, 전남)의 생산 설비가 이용되었다. 다시마, 양파, 알로에, 매실, 케일을 시중(광주광역시 각화동 농산물 도매시장)에서 구입하여 가식 부위를 실험재료로 사용하였다. 각 원료를 깨끗이 씻고, 다시마와 알로에 그리고 케일은 사전 절단작업을 거쳤다. 그리고 모든 원료를 각각 롤러를 이용하여 약 2 mm 이하로 잘게 분쇄하고, 설탕을 총량의 50%(w/w)가 되게 첨가하여 각각 유약을 바르지 않은 항아리(180 L)와 스텐리스강 재질의 용기(200 L)에 넣어 당장하였다. 저장 온도는 20-25°C, 습도는 30±1%를 유지했다. 저장 기간 동안 주 2회 교반하였다. 당장 개시 후 1, 2, 3, 6개월 그리고 12개월째에 75 mL의 각 시료를 무균적으로 채취하여 즉시 실험하였다.
pH는 시료액 20 mL에 대하여 pH meter(Model 520, Orion Research Inc., Boston, MA, USA)를 이용하여 측정하였다. 총산도는 각 시료액 10 g에 증류수 100 mL를 가하여 혼합한 후 0.1N-NaOH 용액으로 pH 8.3이 될 때까지 적정하여 소비된 NaOH 용액의 양(mL)을 젖산 함량(%)으로 나타냈다. 당(설탕, 포도당, 과당)과 유기산(구연산, 주석산, 호박산, 젖산, 초산, 프로피온산)은 HPLC(Shimadzu LC-20AD, Shimadzu Co., Kyoto, Japan)를 이용하여 분석하였다. 즉, 시료액은 Sep-pak® plus C18 cartridges(Waters, Milford, MA, USA)로 정제하여 20 μL를 30°C로 유지된 컬럼(Aminex HPX-87H, Bio.Rad laboratories)에 주입하였다. 이동상으로는 6.5mM H2SO4를 0.5 mL/min 유속으로 사용하였으며, 검출기는 당의 분석에 RI 검출기(Shimadzu RID-10A), 유기산의 분석에는 UV 검출기(Shimadzu SPD-20A, UV 210 nm)를 사용하였다.
일반세균수의 측정은 식품공전에 수록된 PCA 배지를 사용한 표준평판법으로 수행하였다. 젖산생성 세균은 20% 포도당과 1% 탄산칼슘을 첨가한 MRS 한천(Difco Lab., Sparks, MD, USA) 배지를 사용하였다. 먼저 시료를 단계적으로 희석하여 평판 표면에 도말법으로 접종한 후 30°C에서 7-10일간 배양하였다. 그리고 배지 중에 투명 환을 형성하는 모든 집락을 젖산생성 세균으로 선택하였으며, 순수분리를 위하여 3회 계대 배양하였다. 젖산을 생성하는 우점종 세균은 희석 배수를 높인 평판으로부터 선발하였다.
탄수화물 대사에 의한 예비동정은 API 50 CH 키트(BioMerieux, Marcy, l’Etoile, France)를 이용하였으며, 16S rRNA 유전자 염기서열 분석으로 최종적으로 확인하였다. 16S rRNA 유전자의 증폭(Baker 등, 2003)은 콜로니 PCR을 사용하였다. 단일 colony로부터 1 toothpick 채취하여 PCR-premix(iNtRON Biotechnology, Korea)와 혼합한 후, bacteria에 특이적인 primer(8F 5'-AGAGTTTGATCCTGGCTCAG-3'; 1492R 5'-TACGGYTACCTTGTTACGACTT-3')를 사용하여 94°C에서 30초간 변성, 55°C에서 30초간 결합, 72°C에서 50초의 신장 조건으로 22 cycle을 행한 후 최종적으로 72°C에서 10분간 DNA를 증폭시켰다. 증폭된 PCR 산물 크기를 1% agarose gel에서 확인한 후, ㈜Solgent에서 16S rRNA 유전자 염기서열을 분석하였다.
각 미생물의 16S rRNA 염기서열에 대하여 GenBank database(NCBI database)를 통해 기존에 보고된 미생물의 유전자 염기서열과 상동성을 비교하였다. 그리고 각 염기서열을 BioEdit Sequence Alignment Editor program(version 7.0.5; North Carolina State University, Raleigh, NC, www.mbio.ncsu.edu/BioEdit/bioedit.html)의 ClustalW Multiple alignment로 Multiple Sequence Alignment한 후, MEGA 4.0을 사용하여 phylogenetic tree를 작성하였으며, 해당 분석은 neighbor-joining 알고리즘을 사용하였다(Tamura 등, 2007).
농도별(0%, 10%, 20%, 30%, 40% 그리고 50%, w/w)로 설탕을 첨가한 MRS 액체 배지 10 mL에 흡광도 값이 0.4가 될 때까지 배양된 세균 배양액 10 μL를 각각 접종하여 30°C에서 배양하면서 UV-Vis spectrophotometer(S-1100, Scinco, Seoul, Korea)를 이용하여 660 nm에서 24시간 간격으로 4일 동안 측정하였다(Saitou와 Nei, 1987). 그리고 각 미생물이 4일 동안 시료 중에 생육하면서 생산 또는 소비한 당과 유기산의 함량을 HPLC를 이용하여 분석하였다.
결과 및 고찰
식품 재료와 당을 동량으로 당장하면서 기간별 pH는 전체적으로 점점 낮아졌으며, 이 경향은 저장 용기의 종류와는 관계가 없었다(Fig. 1). 매실의 경우, 당장 6개월째에 다른 4종류의 삼출액보다 현저히 낮은 값(2.90-2.99)을 나타내었다. 당장 개시 후 1개월 만에 다시마의 경우(pH 4.55-4.59)를 제외하고, 모두 pH 4 이하가 되었다. 이들 원재료의 pH는 각각 다시마 5.78, 양파 5.84-6.18, 알로에 5.24, 매실 2.73, 케일 5.24로 보고된 바 있다(Bae와 Kim, 2010; Kim 등, 2018; Muñoz 등, 2015; Noisopa 등, 2010; Oancea와 Draghici, 2018). 그러므로, 본 연구에서 설정한 시료 최초 채취일인 당장 1개월째에는 이미 삼출 과정이 상당 부분 진행되었을 것으로 판단되었다. 모든 삼출액의 pH는 당장 6개월 이후에는 안정적으로 낮은 값을 나타냈다. 다시마와 양파의 경우만 서로 다른 통에 저장할 경우, 각각 스텐리스강 용기와 항아리에서 pH가 더 낮았다. 따라서 추후 품질 검사를 바탕으로 하여 시료에 따라 더 적절한 저장 용기의 선택이 가능할 것이다. 이러한 pH의 감소는 시료에 따라 저장 초기에 내삼투압성을 갖고 있으며, 산을 생성하는 미생물이 존재할 가능성이 있음을 시사한다(Son 등, 2005). 그런데 매실 삼출액은 당장 초기부터 pH 3.5에서 생육할 수 있는 젖산균마저 증식하기 어려운 정도의 낮은 pH를 보였다(Song과 Jung, 2006). 일반적으로 강산보다는 해리도가 낮은 유기산이 미생물에게는 더 치명적이라고 알려져 있다(Warnecke와 Gill, 2005). 따라서 저장 12개월째에 접어든 5종의 삼출액은 pH 3.5 정도로서 그 저장성이 우수할 것으로 생각한다.
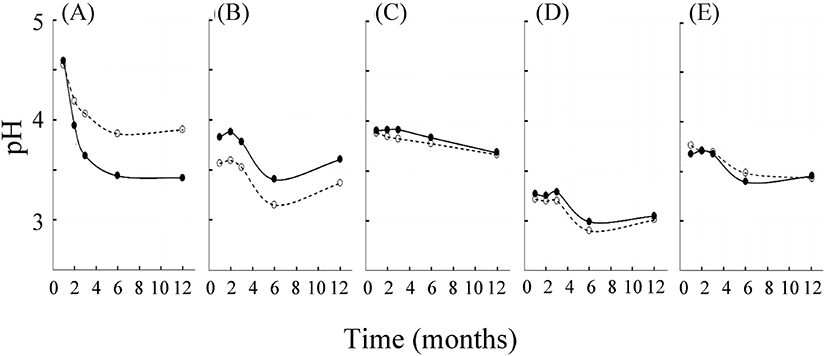
한편, 당장 기간별 총산도의 변화는 1개월 저장한 삼출액들의 경우 산도(%)가 다시마(항아리: 0.36±0.15, 스텐리스강 용기: 0.48±0.27), 양파(0.76±0.28, 0.73±0.08), 알로에(0.83±0.23, 0.75±0.23), 매실(2.77±0.54, 3.29±0.74), 케일(1.02±1.05, 1.49±0.31)로서 pH의 경우와는 달리 12개월까지 등락하는 양상이었으나 유의미한 차이는 아니었다(data not shown).
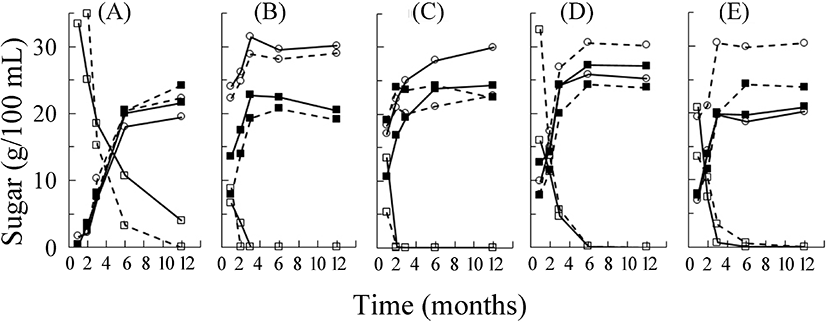
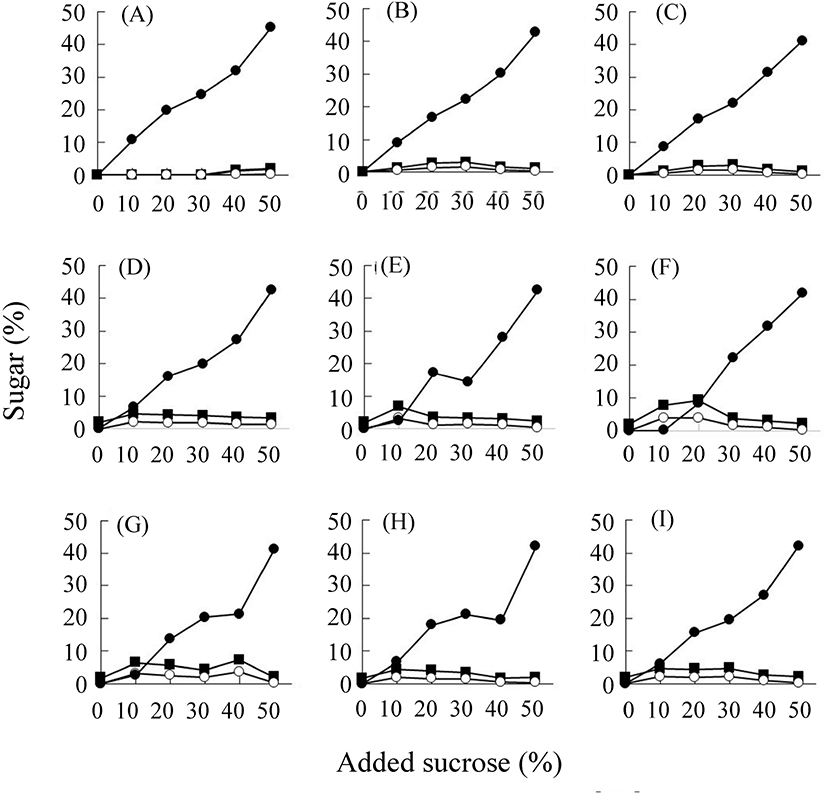
당장 기간에 따라 삼출액 중에는 첨가된 이당류인 설탕의 함량이 감소하는 대신 단당류인 포도당과 과당의 함량은 증가하였다(Fig. 2). 이러한 설탕의 분해 속도는 삼출액 제조 원료에 따라 다름을 알 수 있었다. 저장 2개월째에 다시마가 약 30%의 설탕 농도를 보인 반면, 알로에와 양파는 거의 남아 있지 않은 정도를 보여 빠르게 분해되었음을 알 수 있다. 설탕은 산이나 효소에 의해 그 구성 당인 포도당과 과당으로 가수분해된다. pH 감소 폭이 작은 다시마의 경우만 포도당 농도 증가속도가 현저하게 작은 것으로 보아, 설탕의 분해에 의한 포도당과 과당의 생산이 삼출액 중에서 산의 영향을 받음을 알 수 있다. 대두와 유산발효 대두의 유리당을 분석한 연구에서는 발효 후에 전체적인 유리당의 총량이 현저히 감소하였는데, 이는 발효과정 중 미생물에 의해 당류가 영양원으로 이용되었기 때문으로 보고하였다(Song과 Jung, 2006). 본 실험에서는 설탕, 포도당 그리고 과당의 세 가지 당을 합한 총 당의 함량이 당장 기간의 증가에 따라 크게 변화하지는 않았다. 이는 당류가 미생물에 의해 다량 이용되지 않았음을 시사하며, 설탕의 분해는 미생물 효소 작용보다는 유기산에 의한 산 가수분해임을 보여준다. 그러나 저장 1개월째에 채취한 시료의 pH가 매우 낮은 것으로 보았을 때 1개월 전까지는 미생물이 당류를 분해하여 산을 생성했을 수 있다(Son 등, 2005). 또한, 저장 용기에 따른 변화로는 특히 다시마의 경우, 저장 초기에는 스텐리스강 용기의 경우보다 항아리 용기 중의 설탕 분해가 적었으나, 3개월 이후에는 더 많은 양이 분해되어 용기에 따른 영향이 관찰되었다(Fig. 2). 즉, 이러한 차이가 나는 것은 항아리의 미세기공을 통한 공기의 출입과 관련이 있다고 사료된다(Han 등, 2013).
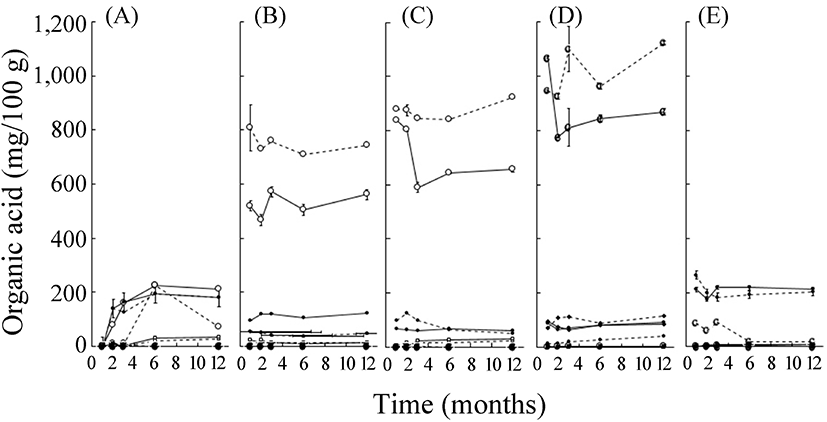
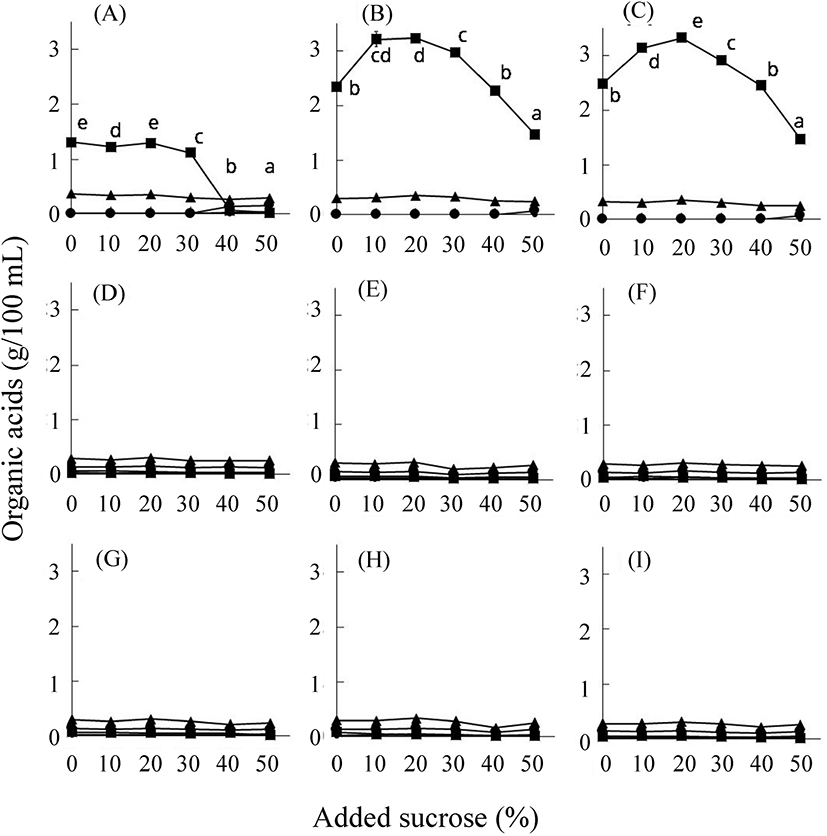
당장 기간에 따른 유기산의 종류 및 함량은 삼출액 제조 원료에 따라 다르게 나타났다(Fig. 3). 다시마의 경우, 구연산, 주석산, 호박산, 젖산, 초산 그리고 프로피온산이 모두 검출되었고, 저장 기간에 따라 감소 또는 증가하는 유기산의 종류가 다름을 확인하였다. 저장 1개월 이후 삼출액 중의 유기산 함량은 매실, 알로에, 양파의 경우 100 g 당 0.8-1 g 정도였다. 이와는 달리 다시마와 케일은 0.2 g의 낮은 값을 보였다. 한편, 다시마, 양파, 알로에는 유기산 중 젖산이 가장 많았으며 이 중에서 양파와 알로에는 항아리를 사용한 경우가 더 많았다(Han 등, 2013). 한편, 매실 삼출액 중에는 구연산이 다량 들어 있었으며, 케일은 전체적인 유기산의 함량은 적으나, 그중에서 초산함량이 가장 많았다. 매실 삼출액 중의 높은 구연산의 함량은 원재료에(Shin, 1995) 기인하나, 케일 중의 초산은 원재료 중에는 함유되지 않아 삼출액 저장 중 생산된다고 할 수 있다(Ayaz 등, 2006).
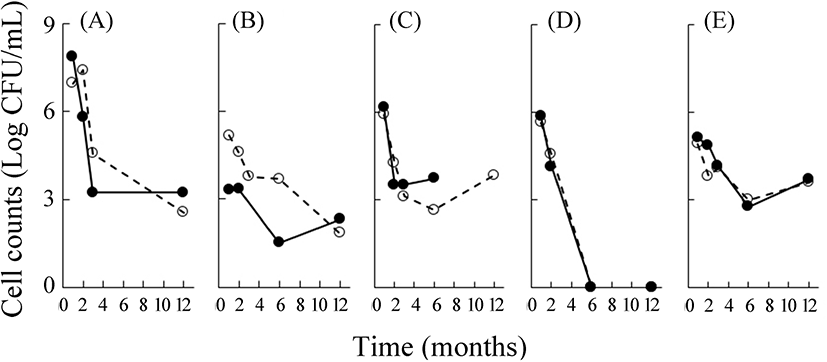
삼출액 중의 일반세균수는 저장 기간이 늘어남에 따라 용기와는 무관하게 감소하였다(Fig. 4). 저장 1개월 후부터는 다시마의 107-108 CFU/mL를 제외하고, 다른 시료들은 103-106 CFU/mL의 범위를 보였다. 모든 시료가 저장 3-6개월까지는 일반세균수가 감소하였고, 이후 그 수준을 유지하였다. 특히 매실의 경우에는 저장 6개월 이후에는 세균이 검출되지 않았다. 이는 매실 중의 구연산이 갖는 세균 사멸 효과 때문일 수도 있다. 따라서 삼출액의 저장 중에는 저장 1개월 이후에는 미생물이 생육하는 양상을 보이지 않았으므로 이 기간의 변화는 일반적인 발효과정이라고 할 수 없으며, 원재료 조직 중의 효소와 미생물의 용균에 따른 균체 효소가 작용한 결과일 수도 있다(Rice와 Bayles, 2008).
20% 포도당과 1% 탄산칼슘을 첨가한 MRS 한천배지를 이용하여 삼출액 중의 젖산생성 세균을 분리하였을 때 5종의 삼출액은 발효 1개월째에는 젖산생성 세균 수가 2×102-2×105 CFU/mL 이하로 나타났다(Fig. 5). 그리고 저장 기간이 지남에 따라 그 수가 점점 줄어들어 12개월째에는 2×101-1×102 CFU/mL 이하가 되었다. 대상 우점종 미생물은 다시마 삼출액에서 발효 2개월째에 2종, 알로에 삼출액에서 발효 1개월째에 3종, 양파 삼출액 저장액에서는 발효 12개월째에 2종, 그리고 케일 삼출액에서는 발효 3개월과 6개월째에 11종이 분리되었으며, 매실 삼출액 저장액에서는 산을 생산하는 세균을 검출할 수 없었다.
매실을 제외한 다시마, 양파, 알로에 그리고 케일 삼출액 저장액에서 산 생성균으로서 선발된 총 18 균주의 염기서열을 BioEdit Sequence Alignment Editor program의 ClustalW Multiple alignment를 이용하여 비교한 결과, 크게는 3개, 세부적으로는 9개로 균주를 구분할 수 있었다(Table 1). 크게 나누었을 때는 SS2-22와 SS2-29, AS1-10, AS1-15, AS1-21과 양파와 케일에서 분리한 나머지 미생물로 나누어졌다. 일단 SS2-22는 나머지 17 균주와 다른 염기서열을 가지고 있었다. SS2-29와 AS1-10, 15, 21은 염기서열 하나만 달랐고, 양파와 케일에서 분리된 13개의 미생물은 각각의 16S rRNA 염.기서열이 작게는 4개, 많게는 9개가 달랐다(data not shown). 이를 동정한 결과, Lactobacillus homohiochii. Lactobacillus plantarum 그리고 Bacillus megaterium으로 나타났으며, 99-100%의 유사성을 나타내었다. SS2-22 균주는 다시마 시료에서 분리되었는데, L. homohiochii LMG 9478T와 가장 높은 염기서열 유사성(% identity 99)을 나타내었다. 또, 다시마에서 분리된 SS2-29는 L. plantarum으로 확인되었다(% identity 100). 알로에 시료 중의 AS1-10, AS1-15, AS1-21은 동정 결과, 모두 L. plantarum인 것으로 나타났다. 양파 삼출액과 케일에서 분리된 13개 균주는 B. megaterium에 속하였는데, 4개의 균주(KJ3-19, KJ6-1, KJ6-2, KS6-2)가 % identity 99이었으며, 그 외는 모두 100%를 나타냈다. B. megaterium은 젖산을 생성할 수는 있지만, 젖산세균으로 분류되지 않고 있다. 또한, 이들이 발견된 케일 삼출액 중의 산 조성도 젖산의 함량이 아주 낮아, 비록 해당 삼출액 중에서 산 생성균으로 발견되었지만 그 역할은 미미한 것으로 보인다.
따라서 산을 생성하는 우점종 세균은 삼출액 원료에 따라 다른 것을 알 수 있다. 즉, 다시마 삼출액 중에는 L. homohiochii와 L. plantarum, 알로에에는 L. plantarum, 양파와 케일의 경우에는 B. megaterium이 우세하였다.
5종의 삼출액으로부터 당장 기간에 따라 선발된 우점종 균주 18종을 30°C에서 배양 및 순수 분리한 후 API system으로 탄소원 이용성을 확인하였다. API 50CHL 배지 및 API 50CHB 배지로 49종의 탄수화물 대사를 조사한 결과는 Table 1과 같다. L. homohiochii로 확인된 SS2-22는 49종의 탄소원 중에 D-포도당과 D-과당만을 이용할 수 있어 다른 분리 미생물과는 확연한 차이가 있었다. 따라서 이 세균은 다시마 삼출액 중에서 설탕이 화학적인 방법으로 분해되어 생산되었거나, 원료 고유의 포도당이나 과당만을 이용하여 생육할 수밖에 없다. L. plantarum으로 확인된 SS2-29, AS1-10, 15, 21은 49종의 탄소원 중 공통적으로 L-arabinose를 비롯한 23종의 탄소원 이용성을 가지고 있었으며, SS2-29만 D-arabitol에 양성반응을 나타냈다. 특히 식물 조직을 구성하는 셀룰로스, 헤미셀룰로스, 펙틴, 키틴질 등 다당류의 분해산물인 D-ribose, D-cellobiose, D-galactose, L-arabinose, N-acetylglucosamine, gluconate 등을 이용하는 능력을 갖고 있어 삼출액 중에서 활발하게 생육 및 발효한 것으로 보인다. B. megaterium으로 확인된 나머지 13 균주는 21-27종의 탄소원을 이용할 수 있으며, 글리세롤, D-xylose 등 11가지 탄소원에 대해서는 그 이용 능력이 균주마다 조금씩 차이가 있었다.
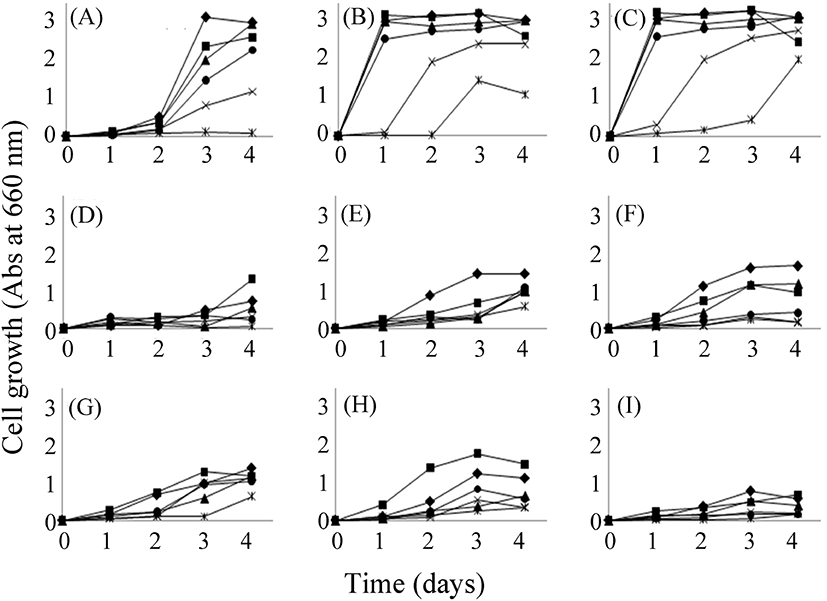
분리 동정된 균주의 발효 특성을 검토하기 위하여 이들의 16S rRNA 염기서열의 phylogenetic tree를 바탕으로 하여 9종으로 압축하였다(data not shown). 이 선발된 균주를 설탕 농도를 달리한 배지에 각 균주를 4일간 생육시킨 후에 그 생육 정도와 당의 함량, 그리고 유기산 함량의 변화를 조사하였다(Fig. 5). 대조구로 사용한 MRS 액체 배지는 발효 전 상태이지만, 소량의 포도당이 함유되어 있었다. 또, 유기산으로서 소량의 구연산, 호박산, 젖산 그리고 초산이 함유된 것으로 조사되었다(data not shown). 케일에서 분리한 B. megaterium의 경우(6종), Lactobacillus sp.보다 낮은 생육도를 보였다. 각 균주는 50% 설탕 함유 배지를 제외한 모든 배지에서 생육하였다(Fig. 5). 특히 분리된 L. plantarum이 잘 생육하여 이 균주가 내삼투압성이 큼을 알 수 있었다(Fig. 5(B), 5(C)). L. homohiochii SS2-22는 설탕 함량이 50%인 경우 4일 동안에는 전혀 생육하지 않았다. 이는 이 미생물이 설탕을 이용하지 못하는 위 실험결과와 일치한다. B. megaterium 균주들은 생육 양상이 서로 유사하였다. 그러나 Lactobacillus 속의 종류들보다는 생육 정도가 낮지만 4일 동안 모든 당 농도에서 꾸준한 생육 양상을 나타냈다. 그리고 설탕의 함량은 첨가한 농도별로 다르게 나타났다(Fig. 6). 당 농도 중 20-40%일 경우 설탕의 분해와 포도당과 과당 증가가 큰 것을 알 수 있다. B. megaterium 균주들에 의한 발효의 경우 포도당과 과당의 생산 능력이 Lactobacillus의 경우보다 컸다. 이들 균주에 의한 주된 유기산 생성은 젖산과 초산의 생성에 있어서 Lactobacillus가 Bacillus보다 10배 정도로 많음을 알 수 있다(Fig. 7). 그러나 L. homohiochii SS2-22는 40% 이상의 설탕 조건에서는 젖산이 거의 생산되지 않았다. 이는 Fig. 2에서 확인된 것처럼 40-50%의 설탕 농도에서 잘 생육하지 못하기 때문으로 보인다. 모든 당 농도 조건에서 젖산을 잘 생산하는 균주는 L. plantarum이었다. 초산의 경우 모든 균주에서 100 mL당 0.3 g 정도로 일정하게 생산됨을 알 수 있었다.
다시마와 양파, 그리고 알로에, 매실 및 케일을 재료로 하여 설탕을 사용한 당 삼출액을 각각 만들고, 이것을 1-12개월간 저장 후 당과 유기산을 분석하였다. 그리고, 산을 생성하는 세균을 분리하여 동정하였다. 저장 기간 동안 모든 삼출액의 pH는 낮아졌다. 설탕은 분해 및 감소되었지만, 포도당과 과당 함량이 늘어나서 전체적으로 당의 함량에는 큰 변화가 없었다. 주요 유기산은 젖산, 구연산, 초산이었으며, 그 양은 삼출액에 따라 달랐다. 또한, 항아리와 스테인리스강 용기를 비교할 때 시료의 종류에 따라 산의 종류별 생성도가 다름이 확인되었다. 이는 삼출액 최종제품의 기호도 향상 조건에 용기의 선택이 고려사항이 될 수 있음을 의미한다. 일반 세균과 젖산 생성 세균의 수는 저장 기간이 길어짐에 따라 감소하였으며, 그 폭은 삼출액의 종류마다 달랐다. 젖산세균으로서 L. homohiochii SS2-22, L. plantarum SS2-29를 분리 및 동정하였으며, 이들 균주는 고농도의 당에서 생육할 수 있으며, 또한 다량의 젖산을 생산함을 알 수 있었다. 이러한 연구 결과는 당을 이용한 삼출액을 제조하는 식품산업을 과학적으로 발전시키는 데 도움이 될 수 있을 것으로 사료된다.
다시마, 양파, 알로에, 매실 그리고 케일의 가식 부분을 대상으로 하여 설탕을 사용한 당장법으로 삼출액을 제조하고, 이를 제조 1개월 후부터 1년까지 항아리와 스테인리스강 용기에 저장하면서 기간별로 당분과 유기산 함량의 변화를 조사하고, 젖산을 생성하는 우점종 세균을 분리 및 동정하였다. 양파와 알로에 삼출액은 젖산 음료에 해당하며, 다시마의 경우 젖산과 초산이 함유되어 있었다. 케일 삼출액 중의 주된 유기산은 초산이었고, 매실액은 구연산 함량이 높았다. 또한, 용기가 삼출물의 종류에 따라 그 삼출물의 조성에도 영향을 미치는 것을 확인하였다. 삼출액의 pH는 전반적으로 낮아졌으며, 일반세균수와 젖산 생성 세균수는 저장 기간이 늘어남에 따라 점점 감소하거나 검출되지 않았다. 세균수는 매실 삼출액의 경우에만 측정되지 않았으며, 모두 18종의 우점종 젖산생성 세균이 확인되었다. 16S rRNA 유전자 염기서열을 확인한 결과, 이들 미생물은 모두 L.homohiochii, L. plantarum 그리고 B. megaterium 속으로 분류되었다. 이 중에서 염기서열이 서로 다른 9종의 미생물을 설탕 첨가 배지에서 4일 동안 배양한 결과, 10% 이하의 조건에서 모두 잘 자랐으며, 특히 Lactobacillus 속은 내당성이 커서 20-50%의 높은 당 농도에서도 생육하였다. 이들은 종에 따라 차이는 있지만, 설탕을 포도당과 과당으로 가수분해하며, 젖산을 발효 생산하였다. 우점종 젖산 생성 세균이 삼출액의 종류에 따라 서로 다름도 확인하였다. 이 연구 결과는 이들 미생물을 스타터로 하여 당장법에 따른 삼출액을 제조하는 등 식품산업에 응용될 수 있을 것이다.