








서 론
대표적인 마늘 가공품인 흑마늘은 첨가물 없이 생마늘을 60-80°C에서 습도를 조절하면서 10일-3개월 정도 숙성시켜서 얻어지는데, 이러한 열처리 숙성 조건 동안에 Maillard 반응과 효소적 반응이 일어나 마늘의 색이 검어지고 향이나 맛, 화학적 조성이 변화된다(Kim 등, 2012). 흑마늘은 외관이나 관능적 특성뿐만 아니라, 이화학적으로도 생마늘과 차별화되는 특성을 가지는데, 열처리 숙성 과정에서 생산되는 Maillard 반응 물질 이외에 페놀화합물, 플라보노이드 등과 같은 항산화성 물질과 더불어 S-allyl cysteine(SAC)과 같은 기능성 성분을 지니게 된다(Kimura 등, 2017). 흑마늘의 대표적인 생리활성 물질로 알려져 있는 SAC는 allyl chloride, allyl bromide, sodium allyl sulfide와 allyl nitrite를 포함하는 allyl halide나 ester가 glutamate 또는 glutathione과 결합하여 얻어진 대사산물로 알려져 있는데, S-allyl glutathione의 glutamate와 glycine 잔기는 γ-glutamyl transpeptidase(γ-GTP)와 dipeptidase에 의해 분해되어 생성된다(Krause 등, 2002). 한편으로 SAC는 γ-GTP가 불활성화되는 열처리 조건에서는 alliin으로부터 직접 생성되기도 한다(Chen 등, 2017). 효소와 비효소적 반응에 의해서 생성된 SAC는 산화 스트레스를 개선하고, 항산화 방어 시스템을 강화하며(Ashafaq 등, 2012), 신경보호(Garcia 등, 2014)와 망막보호(Chao 등, 2014) 효과 및 항염증과 세포자사의 방지 효과(Colin-Gonzalez 등, 2015) 등과 같은 생리기능을 가지는 것으로 알려져 있다.
국내에서 흑마늘과 관련한 연구는 2006년부터 시작되어 흑마늘을 숙성시키는 장치와 숙성 조건 확립, 흑마늘을 이용한 가공식품의 개발, 흑마늘의 다양한 생리활성 규명에 대한 연구 등 다양한 측면에서 연구가 진행되고 있다. 최근 5년간 흑마늘과 관련한 연구로는 방사선 조사시 흑마늘 추출물의 혈구 및 장점막에 대한 보호 효과(Jung 등, 2016) 및 흑마늘을 첨가한 김치소(Lee와 Yoon, 2017), 와인(Ha 등, 2016), 간장(Sim 등, 2016), 식초(Seo 등, 2016)의 제조와 관련한 연구들이 진행되어 있다. 하지만 마늘과 같이 토양에 재배하는 대부분의 작물 등은 기본적으로 토양의 화학성분 조성에 따라 생육 및 성분의 차이를 보이는데, 재배식물의 수량 증가와 관련하여서는 작물의 종류나 재배 지역에 따라 차이는 있지만 품종은 약 50%, 그 외에는 재배기술이나 기상요인 등이 영향을 미치게 된다(Kim과 Ra, 2019b). 따라서 흑마늘의 품질이나 이화학적 특성은 원료 마늘의 특성과 더불어 숙성 온도, 시간 등 가공 조건이 영향을 미치지만, 아직까지 이와 관련하여서는 코끼리 흑마늘과 일반 흑마늘의 항산화활성 비교(Kim 등, 2019)와 동결처리를 통해 흑마늘 숙성 공정을 개선하고자 한 연구(Choi 등, 2017)가 있을 뿐이다.
신품종 마늘인 ‘홍산’은 국제 신품종보호연맹 협약으로 인해 품종보호 작목으로 지정된 마늘의 육종을 위하여 개발된 것으로 2003년 임성자원 ‘8902’와 ‘9209’간의 교배조합으로 얻은 진성 종자 중에서, 최종 선발된 12번째 종자로부터 유래한 마늘구의 영양번식을 통하여 생산력검정시험과 3년간의 지역적응시험을 거쳐 2014년도에 육성되어 품종보호권(제6320호, 2016년)이 등록되었다(Kim과 Ra, 2019a; RDA, 2018). 홍산은 1976년 홍콩에서 중국 가정 지방의 가정백마늘을 도입육종을 위해 들여와 남해, 해남, 제주에서 생산력 검정을 거쳐 우량종을 선발하여 1983년 품종화하면서 명명된 남도마늘(RDA, 2017) 이상으로 월등히 수량이 높고 한지와 난지 모두 재배가 가능하며, 기능성 성분이 높고, 천근성으로 뿌리가 현저하게 적어 수확이 쉬워 생력재배에 알맞다고 알려져 있다(RDA, 2018; RDA, 2019). 홍산 품종의 단점으로는 구의 선단부위에 클로로필로 인해 연한 녹색을 띄는 특징이 있어(NIHHS, 2015) 소비자가 녹변 발생으로 오인 가능성이 높아 개선이 요구되지만 마늘의 외형이 우수하여 흑마늘 제조시 통벌어짐 현상으로 인해 상품성이 떨어지는 난지형 마늘의 단점을 보완할 수 있다.
홍산은 최근에 선발된 품종이라는 특성으로 인하여 재배 외에 식품학적 관점에서 접근한 연구는 식물화학적 구성 및 생리활성(Kim과 Ra, 2019a)에 대한 보고와 남해와 제주, 전주에서 각각 재배한 홍산 마늘의 alliin 함량을 비교한 결과 재배 지역에 따른 통계적인 유의차가 없었으며, 토양의 질소 비율이나 토양 조성 등과 같은 재배환경이 중요한 영향인자라는 보고(Han, 2019)만이 진행되어 있어 관련한 다양한 연구들이 진행될 필요성이 있다.
본 연구에서는 남해군에서 재배한 홍산과 대표적인 재배 품종인 남도마늘로 흑마늘을 각각 제조하여 이들의 이화학적 특성과 더불어 흑마늘로 숙성된 후 이들 성분의 변화를 비교 분석함으로써 가공특성 확인을 위한 기초자료를 확보하고자 하였다.
재료 및 방법
홍산은 농촌진흥청으로부터 분양 받은 것을 남해군에서 재배·수확하여 농가에서 2주 정도 건조한 상태로 출하한 것을 사용하였으며, 남도마늘도 남해군 관내에서 재배되어 홍산과 동일한 과정을 거쳐 건조, 출하된 것을 사용하였다. 흑마늘 제조는 (재)남해마늘연구소 특허(2019)에 따라 45°C에서 시작하여 85°C까지 상승시켰다가 다시 온도를 60°C까지 서서히 내리는 조건에서 항온항습기를 이용하여 12일 동안 숙성하였다. 통마늘 상태의 생마늘을 0일 차 시료로 하고, 2일 간격으로 시료를 채취하여 분석용 시료로 사용하였다.
마늘 시료는 껍질을 제거한 다음 색차계(Ultra scan VIS, Hunter Associates Lab. Inc., Reston, VA, USA)를 이용해 시료의 표면색을 측정하였다. 이 때 표준색판의 L, a, b 값은 각각 99.4, -0.12 및 0.04였으며, 각 숙성 일차별로 10개씩의 시료를 취하여 색도를 측정하였다.
껍질을 제거한 마늘을 7 mm 두께로 슬라이스한 다음 물성측정기(TAXT express, Stable Micro Systems, Godalming, Surrey, UK)를 이용하여 각 시료군당 10개 이상의 시료를 취하여 전단가를 측정하였다. 이 때 분석 조건으로는 Φ 4 mm core를 사용하였고, pre-test speed는 1.0 mm/s, tigger force 50.0 g, test speed 5.0 mm/s, test distance 5.0 mm, test cycle은 1.0이었다.
수분은 껍질을 제거한 마늘을 분쇄기로 분쇄한 후 1 g을 취하여 자동수분측정기(MB45, Ohaus, Zürich, Switzerland)로 측정하였다.
숙성 일차별 마늘을 분쇄한 시료 10 g에 정제수를 가하여 혼합 분쇄한 후 100 mL가 되도록 정용하였다. 이를 충분히 혼합 교반하여 추출한 다음 여과한 시료액을 분석용 시료로 사용하였다. pH와 산도는 여과한 여액 50 mL를 취하여 자동적정기(G20 compact titrator, Mettler toledo, Columbus, OH, USA)를 이용해 동시에 분석하였다. 총당의 함량은 황산페놀법을 이용하여 여과한 마늘 시료액 1 mL에 5% 페놀 용액 1 mL 및 진한 황산 5 mL를 차례로 가하여 30분간 실온에서 정치시킨 후 분광광도계(Libra S 35, Biochrom, Cambridge, England)로 470 nm에서 비색 정량하였다. 표준당으로 맥아당(Sigma-Aldrich Co., St, Louis, MO, USA)을 사용하여 표준검량곡선을 작성하고, 이에 따라 총당의 함량을 산출하였다. 환원당의 함량은 DNS법에 따라 추출 여과한 다음 정제수로 10배 희석한 시료액 1 mL에 DNS시약 3 mL을 가한 후, 끓는 물에서 15분간 중탕 가열한 다음 찬물에서 냉각하여 570 nm에서 흡광도를 측정하고, 포도당(Sigma-Aldrich Co.)을 표준물질로하여 작성한 검량곡선에 따라 정량하였다.
총페놀화합물은 Foiln-Denis 방법(1915)에 따라 시료 무게 대비 10배의 정제수를 가하여 진탕 혼합한 다음 여과한 여액 1 mL에 Foline-Ciocalteau 시약 0.5 mL를 넣고 3분 후 10% Na2CO3 용액 0.5 mL씩을 혼합하여 실온의 암실에서 1시간 정치한 다음 분광광도계(Libra S 35, Biochrom)로 760 nm에서 흡광도를 측정하여 정량하였다. 표준물질로 gallic acid(Sigma-Aldrich Co.)를 사용하여 시료와 동일한 방법으로 분석하여 얻은 검량선으로부터 총페놀화합물의 함량(GAE mg/g)을 산출하였다.
총플라보노이드 함량은 Moreno 등(2000)의 방법에 따라 시료액 1 mL에 10% aluminum nitrate 0.2 mL, 1 M potassium acetate 0.2 mL 및 80% ethanol 4.1 mL를 차례로 가한 후 혼합하여 실온의 암실에서 40분간 정치한 다음 분광광도계(Libra S 35, Biochrom)로 415 nm에서 흡광도를 측정하여 정량하였다. Quercetin(Sigma-Aldrich Co.)을 표준물질로 하여 얻은 검량선으로부터 총플라보노이드 함량(QE mg/100 g)을 계산하였다.
Schwimmer와 Weston(1961)의 방법에 따라 분쇄한 마늘 1 g에 10% trichloroacetic acid 5 mL를 첨가한 다음 균질화하여 1시간 방치한 후 여과지로 여과하였다. 이 여액 1 mL에 0.0125% dinitriphenylhydrazine 1 mL를 가하여 잘 혼합한 다음 37°C에서 10분간 반응시키고, 0.6 N NaOH 용액 5 mL를 가하여 420 nm에서 흡광도를 측정하였다. 표준용액으로 sodium pyruvate를 0.2 μM의 간격으로 농도 조정해 동일한 방법으로 실험하여 얻은 표준검량곡선으로부터 total pyruvate의 함량을 산출하였다.
Thiosulfinate는 Freeman과 Mcbreen(1973)의 방법에 따라 2 mM cysteine이 함유된 pH 7.5의 50 mM N-[2-Hydroxyethyl] piperazine-N'-2-ethane sulfonic acid(HEPES) 용액 0.5 mL와 시료에 일정량의 정제수를 가하여 추출 여과한 여액 0.1 mL를 혼합한 후 50 mM HEPES를 가하여 총 반응용액의 부피를 5 mL로 만든 다음 27°C에서 10분간 반응시켰다. 이 반응액 1 mL를 취하여 50 mM HEPES로 제조한 0.4 mM 5,5'-dithio-bis 2-nitrobenzoic acid(DTNB) 1 mL를 가하여 잘 혼합한 후 다시 27°C에서 10분간 반응시킨 다음 412 nm에서 흡광도를 측정하였다. 50 mM HEPES로 제조한 0.05-0.3 mM의 cysteine 용액을 표준물질로 하여 시료와 동일한 방법에 따라 흡광도를 측정한 결과를 이용해 표준검량곡선을 작성하여 마늘 중의 total thiosulfinate 함량을 산출하였다.
SAC와 γ-GSAC는 동일한 방법으로 전처리한 후 동일한 조건에서 분석하였다. 즉, 시료 5 g에 3차 정제수 45 mL를 가하여 균질화한 후 30분간 sonication하여 추출한 다음 여과지로 여과한 여액을 0.22 μm membrane filter로 재여과하여 HPLC-PDA-MS/MS(TSQ Quantum LC-MS/MS, Thermo scientific, Waltham, MA, USA)로 분석하였다. 분석용 컬럼은 Agilent Zorbax SB-C18(4.6×250 mm, 5 μm, Agilent Technologies, Santa clara, CA, USA)을 사용하였고, 양이온 모드에서 이동상 용매는 0.1% formic acid 수용액과 acetonitrile에 용해한 0.1% formic acid를 시간에 따라 혼합 비율을 조절하였다. SAC(Sigma–Aldrich Co.)와 γ-GSAC (US Pharmacopeia, MD, USA) 표준물질을 시료와 동일한 조건에서 분석하여 머무름 시간을 비교해 확인하였으며, 각각의 표준 검량곡선으로부터 함량을 산출하였다.
결과 및 고찰
홍산과 남도마늘로 흑마늘을 숙성하면서 색의 변화를 분석한 결과는 Fig. 1과 같다. 생마늘에서 홍산과 남도마늘 각각의 명도는 82.0과 83.7로 서로간에 유의적인 차이가 없었으며, 숙성이 시작되면서 급격하게 감소하여 숙성 2일 차에 홍산은 46.0으로 생마늘에 비해 약 44%, 남도마늘은 40.9로 약 51%가 감소하였다. 이후부터는 감소폭이 줄어 완만히 감소하였고, 숙성 2일에서 8일까지는 홍산에서 유의적으로 더 높은 값을 유지하였으나, 숙성 10일 이후부터는 품종에 따른 차이가 없었고, 숙성 12일에 홍산과 남도마늘의 명도는 각각 29.2와 29.9로 낮아졌다.
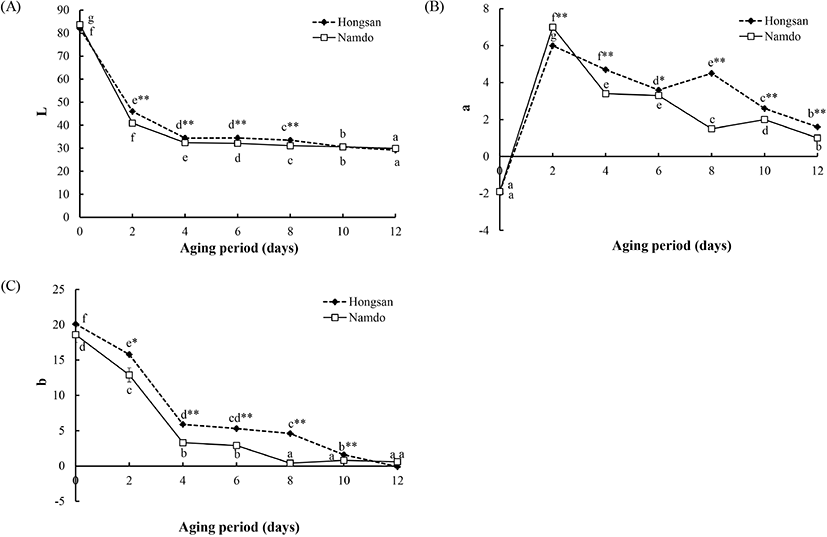
적색도는 생마늘에서는 마늘 품종에 관계없이 동일하게 -1.9였으나, 숙성 2일에 급격히 증가하여 홍산은 6.0, 남도마늘은 7.0으로 최고치를 보였다. 이후 숙성기간의 경과와 더불어 감소하는 경향을 보였는데, 홍산은 숙성 8일에, 남도마늘은 숙성 10일에 각각 한 차례씩 적색도가 증가한 후 다시 감소하였다.
생마늘의 황색도는 홍산에서 20.1, 남도마늘은 18.6이었으며, 서로간에 통계적인 유의차는 없었다. 황색도는 숙성 초기에 급격히 감소하였는데, 숙성 4일에는 숙성 2일에 비해 홍산은 62.7%, 남도마늘은 74.4% 낮아졌다. 이후 홍산은 숙성 4-8일까지는 감소폭이 완만하였다가 다시 유의적으로 감소하였고, 남도마늘은 숙성 8일에 유의적인 감소를 보인 후 숙성 완료시까지 유의적인 차이가 없었다. 숙성 기간 동안 황색도는 홍산이 남도마늘보다 유의적으로 더 높았으나, 숙성이 완료된 숙성 12일에는 서로간에 유의차가 없었다.
흑마늘을 제조하는 과정 중의 색의 변화를 육안으로 관찰하면 대부분의 마늘이 숙성 초기에 급격히 갈변이 일어나고, 숙성 3-4일 정도에 내부까지 진한 갈색으로 변한 다음 숙성이 계속되면서 전체적으로 검은색이 더 진해진다. 색의 분석 결과, 명도, 적색도 및 황색도 모두 숙성 초기에 급격히 감소하고, 이후부터는 서서히 변화를 보이는 것은 이러한 육안 관찰의 결과와 일치하였다.
이상의 색 분석 결과로부터 흑마늘의 숙성 과정에서는 명도보다 적색도나 황색도가 더 큰 영향을 받지만, 숙성이 완료된 이후에는 유의적인 차이가 없어 마늘의 품종은 흑마늘의 색과 관련한 품질에는 영향을 미치지 않음을 알 수 있었다. 흑마늘 숙성 중 껍질의 유무와 숙성 온도를 달리하였을 때 숙성 초기의 명도 값은 서로 상이하지만, 숙성이 완료된 후에는 모두 유사한 범위였다는 보고(Angeles 등, 2016)는 본 연구의 결과와도 일치하였다.
흑마늘 숙성에 따른 경도의 변화는 Fig. 2와 같다. 생마늘의 경도는 홍산에서 1,626.8 g으로 남도마늘(1,068.7 g)에 비해 유의적으로 높았는데, 숙성이 시작되면서 급격히 감소하여 숙성 2일에 홍산은 175.3 g, 남도마늘은 278.4 g으로, 남도마늘에서 유의적으로 더 높았다. 숙성 2일부터 숙성이 완료되는 12일까지 홍산의 경도는 경미한 변화가 있었으나 통계적인 유의차는 없었다. 남도마늘은 숙성 6일에 유의적인 증가 후에 다시 감소하였다가 숙성 10일 이후에는 유의적인 변화가 없었다. 숙성 10일 이후부터는 홍산의 경도가 남도마늘에 비해 유의적으로 더 높았는데, 숙성 12일에 홍산은 143.6 g, 남도마늘의 경도는 70.5 g였다. 이러한 경도의 변화는 흑마늘의 숙성 초기에는 가공 과정에서의 열처리로 인하여 마늘 조직이 물러지지만, 숙성 기간이 경과함에 따라 점차 수분이 감소됨으로써 육질이 단단해지기 때문으로 추정한 보고(Shin 등, 2008)로 미루어 볼 때 숙성 기간 동안 경도의 증감은 수분의 함량 변화와 관련이 있을 것으로 추정된다.
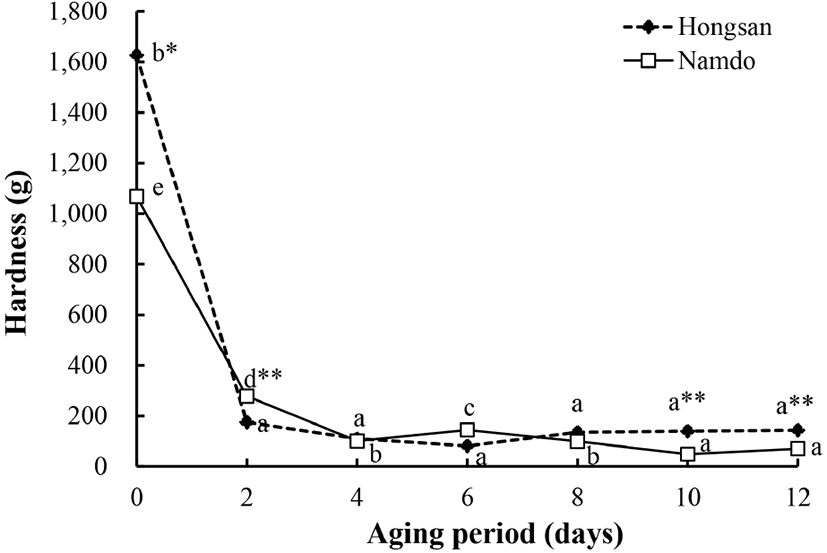
Cha(2012)는 흑마늘의 숙성 초기 경도는 생마늘에 비해 약 85% 낮아지고, 이후 숙성기간이 경과함에 따라 유의적으로 증가하였다고 보고한 바 있으며, Kang 등(2011)은 홍마늘을 숙성시키는 동안 경도는 숙성 초기에 생마늘에 비해 유의적으로 감소한 후 숙성 중기까지는 유의적인 변화가 없다가 숙성 말기에 증가하였다고 보고한 바 있다. 이들 결과는 생마늘에 비해 숙성 초기의 마늘 경도가 급격히 낮아진다는 점에서는 일치하였으나, 숙성 기간에 따른 경도 변화에서는 다소 차이가 있었다. 이와 관련하여 마늘의 숙성 온도가 90°C로 높을 경우 경도는 숙성기간의 경과와 더불어 증가하지만 숙성 온도가 60°C 정도로 낮을 때는 상반된 경향을 보이며, 70°C 정도에서는 숙성 초기에 비해 낮아진 후 변화의 폭이 적다는 Shin 등(2008)의 보고로 미루어 볼 때 기존 연구의 결과들과 본 연구의 결과간의 차이는 마늘의 숙성 온도가 서로 간에 상이하였기 때문으로 생각된다.
홍산과 남도마늘로 각각 흑마늘을 제조하면서 수분, pH 및 산도의 변화를 분석한 결과는 Table 1과 같다. 흑마늘 숙성과정 중 홍산과 남도마늘 모두 수분과 pH는 지속적으로 감소하고, 산도는 증가하는 경향이었다.
수분은 생마늘에서 홍산은 58.14 g/100 g, 남도마늘은 59.68 g/100 g이었는데, 홍산은 숙성 2일 이후에 수분 함량은 50 g/100 g 이하로 낮아진 것에 반해 남도마늘은 숙성 6일까지 50 g/100 g 이상을 유지하였다. 홍산의 수분은 숙성 10일에 37.08 g/100 g으로 낮아졌으나, 남도마늘의 수분은 숙성 12일에 39.84 g/100 g으로 홍산에 비해 유의적으로 더 높았다.
생마늘의 pH는 홍산에서 6.74로 남도마늘(6.26)보다 유의적으로 더 높았으며, 숙성기간 동안에도 남도마늘에 비해 홍산의 pH가 유의적으로 더 높았다. 홍산의 경우 숙성 10일까지도 pH는 5.0 이상이었으며, 숙성 12일에 4.65로 낮아졌으나 남도마늘의 경우 숙성 8일에 pH는 4.75로 낮아진 이후에도 지속적으로 낮아져 숙성 12일에는 4.22였다.
흑마늘 숙성 과정 중 pH의 저하는 열처리 공정 동안 마늘의 세포 파괴로 인한 조직 내부 유기산들의 유리(Yuan 등, 2018), 열처리에 의한 갈변물질의 다량 생산 반응에서 환원당인 aldohexose의 aldehyde기의 산화에 따른 carboxyl기 생성, 당과 염기성 아미노산의 결합에 의한 가용성 염기성 아미노산의 감소 및 산성물질의 생성에 따른 것으로 pH가 낮아지면 갈변반응이 더 촉진된다고 알려져 있다(Choi 등, 1981; Kim 등, 1981).
산도는 숙성 4일까지는 품종에 따른 유의적인 차이가 없었으나, 숙성 6일부터는 남도마늘에서 유의적으로 더 높았다. 홍산의 경우 pH의 변화와 상반되는 경향으로 숙성 2일까지는 유의적인 차이가 없었으며, 숙성 10일에도 0.15 g/100 g으로 증가폭이 적었다가 숙성 12일에 0.24 g/100 g으로 증가하였다. 남도마늘도 숙성 2일까지는 유의적인 변화가 없었으나, 숙성 8일에 0.25 g/100 g으로 증가한 후 숙성 12일에는 0.34 g/100 g으로 홍산보다 약 1.4배 더 높았다.
흑마늘 숙성 중에 일어나는 여러 성분들의 변화는 서로 연계되어 있는데, 당이 분해되면서 acetic acid가 생성되고, hexose나 pentose의 α-, β-dicarbonyl기가 절단되어 단쇄 carboxylic acid가 생성됨으로 인해 formic acid, succinic acid 및 3-hydroxypropionic acid 등이 생성되어 유기산의 함량이 증가하여 산도 증가와 pH 저하가 일어나고, 단백질, 펩타이드 및 다당류의 분해에 영향을 미치는 것으로 보고되어 있다(Liang 등, 2015).
홍산과 남도마늘의 숙성에 따른 환원당 및 총당의 변화를 분석한 결과는 Fig. 3과 같다. 환원당의 함량은 숙성 2일까지는 증가폭이 미미하였으나, 숙성 4일 이후부터는 그 증가폭이 더 커지면서 계속해서 증가하는 경향이었다. 숙성 전 홍산과 남도마늘에서 환원당 함량은 각각 3.34 g/100 g과 3.97 g/100 g으로 남도마늘에서 유의적으로 높았고, 숙성 10일까지는 동일한 경향이었다. 즉, 남도마늘의 경우 숙성 4일에 환원당 함량은 생마늘에 비해 약 3.6배 증가하여 14.16 g/100 g이었고, 숙성 8일 이후부터는 22.34 g/100 g 이상이었으나, 홍산의 경우 숙성 10일에서 13.08 g/100 g이었고, 숙성이 완료되는 숙성 12일에야 24.68 g/100 g으로 증가하여 남도마늘과 유의적인 함량 차이가 없었다.
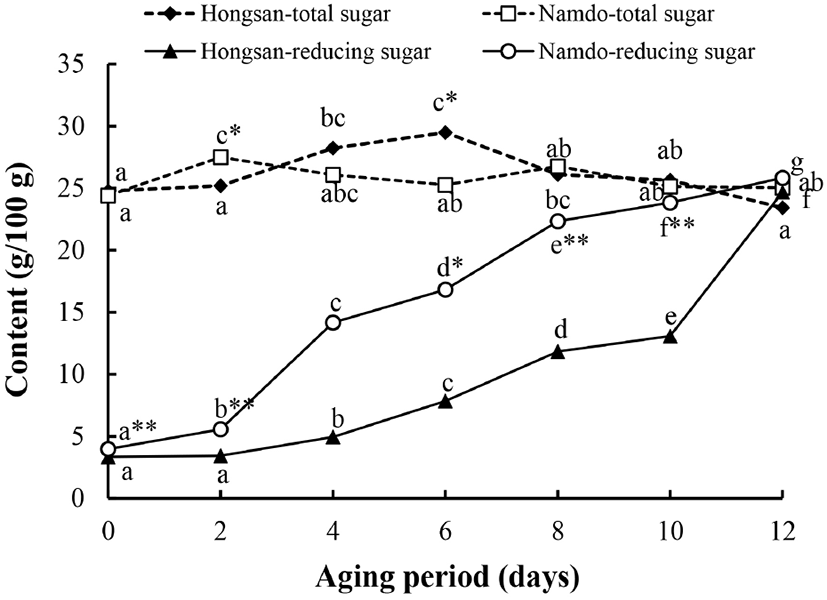
총당의 함량은 홍산에서 23.42-28.23 g/100 g, 남도마늘에서는 24.40-27.49 g/100 g의 범위에서 숙성기간의 경과와 더불어 불규칙한 증감을 보였는데, 생마늘과 숙성 완료된 흑마늘에서 함량은 유의적인 차이가 없었다.
흑마늘 숙성 중 당류의 함량 변화와 관련하여 다당류의 분해는 산성과 고온 조건하에서 일어나는데, 세포벽의 다당체가 분해되면서 조직이 부드러워지고, 흑마늘의 질감이 검과 같이 변화하게 되며, 열처리 동안 pH의 저하는 sucrose가 glucose나 fructose로 분해되는 것을 촉진함으로써 흑마늘 숙성 동안 이들 당류의 함량이 증가한다고 보고되어 있다(Liang 등, 2015).
한편, 환원당의 생성 속도와 관련하여 흑마늘 숙성 중 환원당의 함량은 마늘 내 다당류가 분해되어 생성되는 양과 Maillard 반응 동안 소비되는 양에 의해 영향을 받는데, 70°C 이하의 온도에서는 환원당의 생성속도가 Maillard 반응 등에 소비되는 속도보다 빨라서 그 함량이 지속적으로 증가하지만 80°C 이상에서는 숙성 초기는 환원당의 축적율이 소비량을 능가하여 그 함량이 증가하지만 고온으로 인해 Maillard 반응 속도가 빨라짐에 따라 숙성 후기에는 그 함량이 오히려 감소한다고 보고되어 있다(Zhang 등, 2015).
마늘 중에는 건물량으로 약 75%의 다당류가 함유되어 있으며, 이들의 주성분은 fructan으로(Baumgartner 등, 2000), fructose와 glucose가 14:1 정도의 비율로 혼합되어 있다(Chen 등, 2013). Fructan은 흑마늘로 제조되는 열처리 공정을 거친 후에 84.79%가 감소하며, 이와는 상반되게 fructose 함량은 500% 이상 증가하며, glucose와 sucrose 함량에는 큰 차이가 없다는 보고가 있다(Yuan 등, 2018).
이상의 보고들과 비교하여 볼 때 본 연구의 결과에서는 상대적으로 숙성 온도가 높은 초기에 환원당의 생성량이 증가하였고, 이후 숙성 온도가 낮아지면서 갈변반응에 소비되는 환원당의 함량보다 다당류의 분해로 인해 생성되는 양이 더 많아져서 환원당의 함량은 증가하는 경향을 보인 것으로 생각된다. 또한, 다당류의 분해가 유도되지만 상대적으로 유리되는 가용성 당류의 함량 또한 증가함으로 인해 총당의 함량 변화에는 크게 영향을 미치지 않은 것으로 생각된다.
흑마늘 숙성 과정 중 총페놀화합물 및 총플라보노이드는 숙성 기간의 경과와 더불어 점차 증가하는 경향이었다(Table 2). 총페놀화합물의 함량은 홍산과 남도마늘의 변화 경향이 서로 차이가 있었다. 즉, 홍산에서는 생마늘과 숙성 2일 차 마늘간에 유의적인 함량 변화가 없었고, 숙성 10일까지도 큰 함량의 증가가 없어 생마늘에 비해 약 9% 증가하였다가 숙성 12일에 4.87 GAE mg/g이었다. 남도마늘의 경우, 생마늘에 비해 숙성 6일에는 약 16%가 증가하여 4.24 GAE mg/g이었고, 숙성 12일에는 약 1.8배가 증가하여 6.53 GAE mg/g이었다.
총플라보노이드 함량도 총페놀화합물과 유사한 경향으로 증가하였는데, 홍산은 숙성 4일에 생마늘에 비해 약 4.5배가 증가한 7.17 QE mg/100 g이었고, 숙성 8일까지는 유의적인 차이가 없다가 숙성 10일에 10.98 QE mg/100 g으로 증가하였으며, 숙성 12일에는 14.00 QE mg/100 g이었다. 남도마늘의 총플라보노이드 함량은 홍산에 비해 유의적으로 더 높았으며, 생마늘에 비해 숙성 4일에 약 3.3배 증가하여 9.87 QE mg/100 g이었고, 숙성 10일에는 22.73 QE mg/100 g, 숙성 12일에는 34.16 QE mg/100 g으로 최종 숙성 완료된 후에는 홍산에 비해 약 2.44배 더 높았다.
코끼리 마늘과 일반 마늘로 각각 흑마늘을 숙성한 후 총페놀화합물과 총플라보노이드 함량을 측정한 결과에서도 본 연구와 유사하게 원료 생마늘에서는 총페놀화합물의 함량에 유의적인 차이가 없었으나, 숙성 후에는 코끼리 흑마늘에서 그 함량이 유의적으로 더 높았고, 플라보노이드는 생마늘의 경우 코끼리 마늘에서 유의적으로 함량이 더 높았으며, 숙성 후에도 이러한 경향이 유지되었다고 보고되어 있다(Kim 등, 2019).
흑마늘 숙성 중 총페놀화합물의 함량은 일반적으로 증가하지만, 그 증가 폭은 흑마늘의 숙성 조건에 따라서 서로 상이하여 78°C에서 14일간 숙성할 때보다 75°C에서 21일간 숙성할 때 증가량이 더 높으며, 생마늘 대비 통마늘로 숙성하였을 때는 약 3배, 깐마늘로 숙성한 경우는 약 6배 정도 증가한다고 보고되어 있다(Angeles 등, 2016).
마늘의 함황화합물이나 페놀화합물 및 플라보노이드와 같은 구성 성분들은 품종과 여러 재배 조건 등에 영향을 받지만, 재배 조건보다는 품종이 더 유의적인 영향을 미친다는 Martins 등(2016)의 보고로 미루어 볼 때 본 연구 결과에서 플라보노이드의 함량 차이는 품종의 특성이 영향을 미쳤을 것으로 생각된다. 총페놀화합물의 경우는 품종에 따른 함량 차이보다는 세포벽의 파괴에 따른 페놀화합물의 유리 정도와 열처리에 따른 성분의 분해 속도 차이 등이 더 큰 영향을 미치기 때문에 숙성 정도가 경과함에 따라 함량 차이가 유도된 것으로 추정된다.
홍산과 남도마늘 품종으로 각각 흑마늘을 제조하는 동안 pyruvate와 thiosulfinate 함량의 변화는 Fig. 4와 같이 유사한 경향으로 감소하였다. Pyruvate 함량은 마늘의 품종에 따른 유의적인 차이 없이 생마늘에서 홍산은 10.07 mg/g, 남도마늘은 9.83 mg/g이던 것이 숙성 기간과 더불어 유의적으로 감소하여 숙성 12일에는 각각 3.66 mg/g과 3.69 mg/g으로 차이가 없었다.
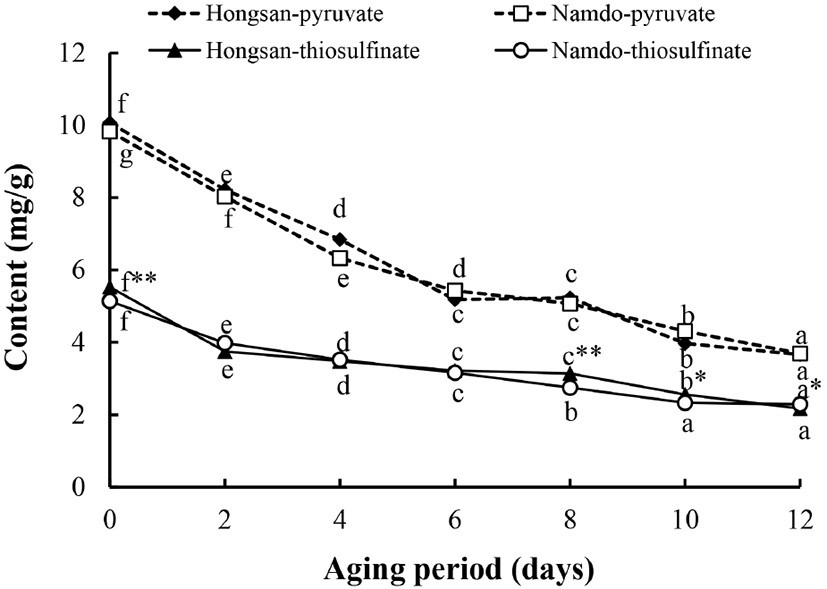
Thiosulfinate는 숙성 전 홍산과 남도마늘에서 각각 5.53 mg/g과 5.14 mg/g으로 홍산에서 유의적으로 높은 함량이었으며, 숙성 2일에 각각 32.3%와 22.6%씩 감소하여 그 감소폭이 가장 컸다. 이후에도 1.7-18.5% 범위에서 점진적으로 감소하는 경향이었으며 숙성 완료 후에 홍산은 2.17 mg/g, 남도마늘은 2.29 mg/g이었다.
Kang(2016)은 흑마늘 숙성을 5단계로 나누어 총 thiosulfinates와 pyruvate 함량을 분석한 결과, thiosulfinates는 숙성 초기 급감한 후 서서히 감소하였는데 이는 Maillard 반응과 SAC 및 S-allylmercapto-cystein(SAMC)이나 다른 물질로 변화하기 때문으로 추정하였으며, pyruvate는 숙성 초기 크게 증가한 이후 감소하는 경향을 보여 흑마늘에서 마늘 냄새 감소에 기여한다고 보고한 바 있는데, 이는 본 연구의 결과와 유사한 경향이었다.
이와는 상반되게 자이언트 마늘과 일반 마늘 각각으로 흑마늘을 만들었을 때 생마늘에 비해 pyruvate 함량은 증가하였으며, 자이언트 마늘로 제조한 흑마늘에서 유의적으로 함량이 더 높았다는 보고(Kim 등, 2015)도 있다. 시판 흑마늘 중의 pyruvate와 thiosulfinate 함량은 서로 비례적으로 존재하며, 마늘의 총 thiosulfinate 함량은 숙성 온도에 따른 큰 차이는 보이는 것으로 보고되어 있다(Cha 등, 2012). 흑마늘 중의 pyruvate와 thiosulfinate 함량에 대한 본 연구결과 및 기존 연구결과들 간의 차이와 관련하여 Lee 등(2010)은 숙성 온도와 기간, 그리고 숙성에 따른 수분 함량 변화로 인해 상대적인 고형분 함량의 차이, 고온에 의한 당의 분해산물 등이 영향을 미칠 것으로 추정하였다.
홍산과 남도마늘의 흑마늘 숙성 중 SAC 및 γ-GSAC 함량을 분석한 결과는 Table 3과 같다. 홍산의 SAC 함량은 생시료일 때 49.57 mg/100 g이던 것이 숙성 2일에 약 3.8배가 증가하여 188.54 mg/100 g이었다. 숙성 4일에는 236.62 mg/100 g으로 증가한 후 숙성 6일까지는 유의적인 변화 없이 유지되었다가 그 이후부터는 다시 점차 감소하여 숙성 12일에는 142.18 mg/100 g이었다. 남도마늘의 SAC 함량은 생마늘에서 51.17 mg/100 g으로 홍산과 유의적인 차이가 없었는데, 숙성 2일에는 157.94 mg/100 g으로 약 3배가 증가하여 최고치였고, 그 이후부터는 숙성기간과 더불어 감소하여 숙성 10일에 77.75 mg/100 g으로 최고치 대비 잔존율은 약 49%에 불과하였다. 숙성 4일부터는 홍산의 SAC 함량이 남도마늘에 비해 유의적으로 높았는데, 숙성 12일에 남도마늘의 SAC 함량은 60.79 mg/100 g으로 홍산의 42.8%에 불과하였다.
본 연구의 결과, 생마늘 대비 최종 숙성 완료된 흑마늘에서 SAC 함량은 홍산은 약 2.9배, 남도마늘은 약 1.2배 더 높았다. 이는 마늘 숙성 전후의 SAC 함량과 관련하여 생마늘 대비 건물량으로 약 5.6배가 증가하였다는 보고(Bae 등, 2014)와 숙성 완료 후 약 6배가 증가하였다는 보고(Hanum 등, 1995), 생마늘 착즙액 대비 흑마늘 착즙액에서는 약 3.3배가 증가하였다는 보고(Kim 등, 2017) 및 생마늘 대비 흑마늘에서 약 2.2배가 증가하였다는 보고(Kim 등, 2012) 등과 비교해 볼 때 증가하는 경향은 일치하였으나 그 절대값은 차이가 있었다. 이러한 차이는 각 연구마다 흑마늘의 숙성 조건이 서로 상이하고, 원료 마늘 중의 SAC 함량 및 보관상태 등이 상이하기 때문으로 추정되는데, Kim 등(2012)은 생마늘을 4°C의 저온에서 1개월 저장하였을 때 SAC 함량은 약 8배 정도 증가하였음을 보고한 바 있다.
γ-GSAC 함량은 홍산과 남도마늘이 유사한 변화의 경향을 보여 생마늘에 비해 유의적으로 증가하여 숙성 2일 차에 최고치였으며, 그 이후부터는 다시 감소하였다. 숙성 전 생마늘에서 γ-GSAC 함량은 홍산과 남도마늘에서 각각 230.86 mg/100 g과 106.92 mg/100 g으로 홍산에서 약 2.2배 더 높은 함량이었고, 숙성이 완료된 12일 차에는 각각 94.72 mg/100 g과 15.81 mg/100 g으로 잔존율은 각각 41%와 14.8%였다.
흑마늘의 숙성 동안 γ-GSAC는 γ-GTP에 의해 분해되어 SAC로 전환되는데, 마늘 중의 수분은 γ-GTP에 의한 가수분해를 도와주어 γ-GSAC로부터 SAC의 생성을 증가시키지만, 고온에서 마늘을 가열하게 되면 수분의 손실이 일어나면서 γ-GTP의 활성도 낮아지게 되므로 γ-GSAC로부터 SAC의 생성 반응은 아직은 알려지지 않은 여러 요인들이 관여한다는 Bae 등(2014)의 보고가 있다.