
1. Introduction
Edible insects are increasingly explored as ingredients in food systems, yet their broader adoption depends on robust evidence of safety and consistent quality. Risk assessments and supply-chain analyses emphasize that insect-based materials may pose biological and chemical hazards originating from production substrates and processing conditions, and that standardization is essential to reduce variability and strengthen consumer trust (Fuso et al., 2024; Liceaga et al., 2022; Traynor et al., 2024). For food-oriented applications, pre-processing steps that influence hygiene, chemical safety, and product quality warrant systematic evaluation (Acosta-Estrada et al., 2021; Fraqueza and Patarata, 2017; Kooh et al., 2020).
Larvae of the white-spotted flower chafer, Protaetia brevitarsis, have been investigated as an insect resource for food applications in Korea (Kim et al., 2024; Park and Kim, 2025; Park et al., 2024; Park et al., 2025b). Previous research has proposed pre-treatment conditions for P. brevitarsis larvae intended for use as food ingredients, highlighting the importance of defining processing workflows before further conversion into powders or other forms (Kim et al., 2022; Kim et al., 2025). Despite this progress, key pre-processing control points that can affect both yield and product quality require additional evidence, particularly under practical farming and processing contexts.
A central pre-processing step for many edible insects is gut evacuation, often termed depuration or fasting, which is applied to reduce intestinal contents prior to downstream processing (Ahn et al., 2015; Chung et al., 2013; Mancini et al., 2020). Because insects are frequently processed and consumed as whole-body materials, residual gut contents can contribute to hygienic concerns and sensory issues (Baigts-Allende and Stathopoulos, 2023; Cappelli et al., 2020; Kooh et al., 2019; Zhang et al., 2024). Feed withdrawal has been used to promote fecal clearance in farmed insects; for example, in the edible cricket Gryllus bimaculatus, fecal output decreased substantially after a short fasting period, supporting depuration as a practical measure to reduce gut contents (Ahn et al., 2015; Kwon et al., 2013; Park et al., 2025a). At the same time, prolonged fasting can impose physiological stress and reduce harvest yield through weight loss or mortality, indicating a trade-off between gut clearance and production efficiency (Egonyu et al., 2025; Han et al., 2024). Therefore, depuration strategies that preserve gut-evacuation benefits while minimizing production losses are relevant to food-grade insect processing.
Diet and processing conditions are well-recognized sources of variability in insect composition (Cacchiarelli et al., 2022; Megido et al., 2017; Selaledi and Mabelebele, 2021). Reviews on edible insects have shown that macronutrient composition, lipid composition, and overall acceptability can vary with species, developmental stage, diet, and processing method (Kouřimská and Adámková, 2016; Meyer-Rochow et al., 2021; Oonincx and Finke, 2021). For P. brevitarsis, changes in the type of feed provided prior to processing have been associated with measurable shifts in nutritional composition, indicating that the pre-harvest period can influence final material characteristics (Ham et al., 2021; Noh et al., 2015). From a safety perspective, chemical contaminants such as heavy metals are relevant hazards for insect-based foods, and empirical studies in farmed insects indicate that diet can influence heavy-metal levels in larvae (Gori et al., 2025; Meyer et al., 2021). Heavy-metal accumulation has also been discussed in relation to insect substrates and feeding conditions, supporting the need for monitoring when alternative ingredients are introduced (EFSA Scientific Committee, 2015; Schrögel and Wätjen, 2019).
Sensory quality is another practical barrier to consumer acceptance of insect-based foods. Studies on beetle larvae have shown that processing interventions can reduce characteristic off-odors associated with specific volatile compounds, underscoring the value of volatile profiling for evaluating strategies designed to improve acceptability (Kim et al., 2021). Because odor-related attributes are often driven by complex mixtures of compounds, multivariate approaches can help interpret treatment-dependent shifts in overall volatile patterns (Baldovini and Chaintreau, 2020; Lubes and Goodarzi, 2017).
In this context, we evaluated whether formulated depuration diets provided during the gut-evacuation step could serve as a controllable pre-processing strategy to modulate the nutritional quality, chemical safety, and volatile profiles of P. brevitarsis larvae. Three formulated depuration diets composed of wheat bran, eggshell powder, and brewer’s spent grain at graded proportions (FD1-FD3) were compared with a glutinous rice flour-based control reflecting common practice. We assessed treatment-dependent differences in nutritional composition (proximate composition and fatty acid/mineral profiles), measured selected heavy metals relevant to food safety, and summarized multivariate variation in volatile profiles using principal component analysis (PCA).
2. Materials and methods
Eggshells used to prepare the formulated diets were immersed in water and washed to remove foreign materials. After washing, excess surface water was removed at room temperature until no visible moisture remained. The eggshells were then dried in a hot-air dryer at 105°C for 2 h to remove residual moisture and sterilize the material. The dried eggshells were ground into a powder and used as an ingredient. Wheat bran and brewer’s spent grain were also dried to reduce moisture. All ingredients were weighed according to the composition shown in Table 1 and mixed thoroughly to obtain a uniform formulated diet for the fasting process.
| Ingredient (%) | Control | Formulated Diet | ||
|---|---|---|---|---|
| GRP1),2) | FD13) | FD2 | FD3 | |
| Glutinous rice powder | 1004) | - | - | - |
| Wheat bran | -5) | 47 | 73 | 90 |
| Eggshell powder | - | 3 | 2 | 1 |
| Brewer’s spent grain | - | 50 | 25 | 9 |
GRP, larvae provided with the control diet (glutinous rice powder) during depuration; FD1-FD3, larvae provided with formulated diets 1-3 during depuration.
P. brevitarsis larvae were reared in the insect-rearing facility of the Industrial Insect Division, National Institute of Agricultural Sciences, Rural Development Administration (RDA, Korea). Rearing conditions were maintained at 25±1°C, 50-60% relative humidity, and a 16L:8D photoperiod. Fermented oak sawdust (Healing Bugs, Gimje, Korea) was used as the rearing substrate/feed. Only late third-instar larvae meeting the harvest body-mass criterion (approximately 2.5 g per individual) were selected (Park and Kim, 2025).
For gut-content removal as a pretreatment, larvae were provided with the control diet (glutinous rice powder, GRP) or one of three formulated diets for 48 h. Each diet was spread to a thickness of 1-2 cm in plastic rearing containers (54×40.5×18.5 cm; length×width×height). Two hundred larvae were placed in each container, and two independent replicate containers were prepared per treatment group under the same rearing conditions. The rearing container was defined as the biological experimental unit (n=2 containers per treatment). After feeding, the diet was removed, and larvae were fasted for 2 days (48 h) with adequate ventilation under a dry environment.
After fasting, larvae were washed three times with water to remove surface contaminants and gently blotted on filter paper to remove excess surface water. Washed larvae were rapidly frozen at -70°C for at least 24 h using an ultra-low temperature freezer (NIHON Freezer, Tokyo, Japan). Frozen samples were then freeze-dried for 65±5 h using a freeze dryer (Illshin Biobase, Dongducheon, Korea). The dried larvae were ground using a 100-mesh grinder (Korea Industry, Siheung, Korea), and the resulting powders were stored at -80°C until analysis.
The proximate composition of P. brevitarsis larvae was determined according to the Korean Food Code of the Ministry of Food and Drug Safety (MFDS, Korea) and Association of Official Analytical Chemists (AOAC) methods. Moisture content was measured by atmospheric-pressure oven drying at 105°C to constant weight. Crude ash content was determined by dry ashing in a muffle furnace at 550°C. Crude protein content was determined using the micro-Kjeldahl method after digestion with a catalyst mixture (protein digestion accelerator) and concentrated sulfuric acid. Total nitrogen was quantified using an automated Kjeldahl analyzer (Kjeltec 8400, GRPS Analytical, Hillerød, Denmark), and crude protein was calculated using a nitrogen-to-protein conversion factor of 6.25. Crude fat content was determined using a Soxhlet apparatus with diethyl ether as the solvent, and the extracted residue was quantified gravimetrically after solvent removal. Crude fiber was determined using a filter bag filtration method. Carbohydrate content was calculated as 100 − (moisture + crude protein + crude fat + crude ash). Results were expressed as weight percentages (% w/w), as specified.
Fatty acid composition was determined according to the Korean Food Code using a Folch-type solvent extraction method, conversion to fatty acid methyl esters (FAMEs), and subsequent GC analysis. Briefly, 50 g of homogenized larval sample was extracted with 250 mL chloroform:methanol (2:1, v/v) using a homogenizer operated at 3,000 rpm. The extract was dehydrated over anhydrous sodium sulfate and concentrated in a water bath. The lipid fraction was saponified with 1 mL of 0.5 N NaOH at 100°C for 20 min and cooled at room temperature for 30 min, then methylated by adding 2 mL boron trifluoride reagent, followed by an additional 20-min heating step and a 30-min cooling period. Phase separation was induced by adding 1 mL n-heptane and 8 mL saturated NaCl solution, and the upper organic layer containing FAMEs was collected. FAMEs were analyzed using GC (US/HP 6890, Agilent Technologies, Seoul, Korea) equipped with an SP-2560 silica capillary column (100 m×0.25 mm i.d., 0.20 μm). The inlet temperature was set to 225°C, and the sample injection volume was 1 μL.
Mineral contents were analyzed according to the Korean Food Code. Dried larval powder (50 mg) was subjected to dry ashing via sequential drying and carbonization, followed by complete ashing at 450-550°C in a muffle furnace. The resulting ash was dissolved in hydrochloric acid and filtered through a glass fiber filter. Elemental concentrations were quantified using inductively coupled plasma-optical emission spectrometry (ICP-OES; Horiba, Kyoto, Japan). Molybdenum (Mo) and selenium (Se) were analyzed after dry-digestion pretreatment. The chlorine (Cl) content was determined by ashing 1 g of a salt-containing sample; the ash was dissolved in water, filtered, and titrated with silver nitrate using potassium chromate as an indicator.
Heavy metal contents were determined according to the Korean Food Code. Samples were dry-ashed at 600°C for >2 h in an electric furnace. After cooling to room temperature, the ash was dissolved in 1:1 (v/v) hydrochloric acid and allowed to stand for 18 h. The solution was filtered through Whatman No. 6 filter paper (Whatman International Co., Maidstone, UK). Lead (Pb), cadmium (Cd), and arsenic (As) were quantified using ICP-OES (Horiba, Kyoto, Japan) at 220.35, 226.50, and 193.70 nm, respectively.
Electronic nose (E-nose) analysis was performed following the methods described by Kang et al. (2020) and Oh et al. (2025) with some modifications. For multicomponent odor profiling, 2.5 g of finely ground sample was mixed with 5 mL of triple-distilled water and stirred. The mixture was transferred to a 20-mL vial (Ls-Phs-Psck GmbH, Langerwehe, Germany), and headspace volatile compounds were generated by incubating the vial at 40°C for 20 min with agitation at 500 rpm. Volatile profiles were analyzed using an E-nose system (Alpha MOS, Toulouse, France) equipped with two columns (MTX-5 and MTX-1701) and a flame ionization detector (FID). The column oven temperature program was as follows: 50°C for 2 s, increased at 1°C/s to 80°C, then increased at 3°C/s to 250°C and held for 21 s. Injection was performed using a syringe-type injector at 25°C with a column head pressure of 1.0 psi. During analysis, injector pressure and detector pressure were set to 1.0 and 39.0 psi, respectively. Volatile compounds were tentatively identified based on retention time and the instrument library, and peak-area data were used for subsequent analyses. PCA was performed using Alpha MOS software to visualize treatment-dependent differences in overall volatile profiles, and discrimination power (DP) values were used to identify marker compounds contributing to sample differentiation.
Results are presented as mean±standard deviation. Statistical comparisons among the four treatment groups (GRP, FD1, FD2, and FD3) were performed using one-way analysis of variance (ANOVA). When significant treatment effects were detected, Tukey’s honestly significant difference (HSD) test was applied for multiple comparisons. Statistical analyses were performed using PASW Statistics 22.0 for Windows (IBM Corp., Chicago, IL, USA), and differences were considered significant at p>0.05. Electronic-nose data were further analyzed using Alpha MOS software (Alpha MOS, Toulouse, France). PCA was applied to the peak-area data to compare overall volatile patterns among treatments, and score and loading plots were used for visualization. Compounds with DP values >0.90 were regarded as major discriminant volatiles.
3. Results and discussion
Proximate composition was determined in whole-larvae powders collected after a gut-emptying pretreatment, comparing the control group (GRP) with larvae subjected to formulated-diet treatments (FD1-FD3) (Table 2). Crude protein content was 53.2±2.9% in GRP and ranged from 55.6±0.7% to 57.4± 0.7% in the FD groups. Although FD1 and FD2 showed numerically higher crude protein than GRP, Tukey’s HSD test did not detect significant differences among treatment groups for crude protein or any other variables listed in Table 2 (all NS).
GRP, larvae provided with the control diet (glutinous rice powder) during depuration (control group); FD1-FD3, larvae provided with formulated diets 1-3 during depuration (formulated-diet groups).
Crude fat was numerically lower in FD1-FD3 (10.8-11.9%) than in GRP (12.5±1.0%), whereas crude ash was numerically higher in FD1-FD3 (8.6-9.9%) compared with GRP (7.5±1.2%). Dietary fiber ranged from 16.5±0.1% to 18.7±0.0%, and carbohydrate ranged from 20.9±2.3% to 26.3±1.4% across the treatment groups. These numerical differences were not statistically significant under the present experimental design.
Collectively, the formulated-diet pretreatment should be interpreted as producing numerical tendencies in proximate composition, dietary fiber, and carbohydrate rather than statistically supported compositional shifts. Such diet-dependent variability in insect proximate composition is well documented; species, developmental stage, feeding substrate, and processing conditions—particularly the feeding substrate—can influence crude protein and crude fat levels (Janssen et al., 2017; Oonincx et al., 2015; Rumpold and Schl, 2013). However, interpretation of short-term pretreatment effects requires caution because the current analysis was performed on whole larvae, including the gastrointestinal tract. Consequently, observed numerical shifts may reflect not only changes in larval tissues but also differences in residual gut contents (e.g., feed residues and frass) remaining at sampling.
To disentangle these effects, future work should (1) quantify the residual mass of gut contents before and after pretreatment and (2) evaluate gut-emptying efficiency metrics (e.g., frass output and gut-content weight) alongside proximate composition, thereby clarifying the pretreatment’s role as a controllable process variable affecting analytical outcomes (Kipkoech, 2023; Yoon et al., 2020).
Thirteen individual fatty acids were quantified (Fig. 1), and oleic acid was consistently the predominant fatty acid across all groups. Oleic acid was highest in FD3 larvae (11.275 g/100 g), followed by FD2 (7.500 g/100 g), GRP (7.050 g/100 g), and FD1 (6.227 g/100 g). Tukey’s HSD grouping indicated that FD3 was significantly higher than FD2, GRP, and FD1; FD2 was significantly higher than FD1; and GRP did not differ significantly from FD1 or FD2.
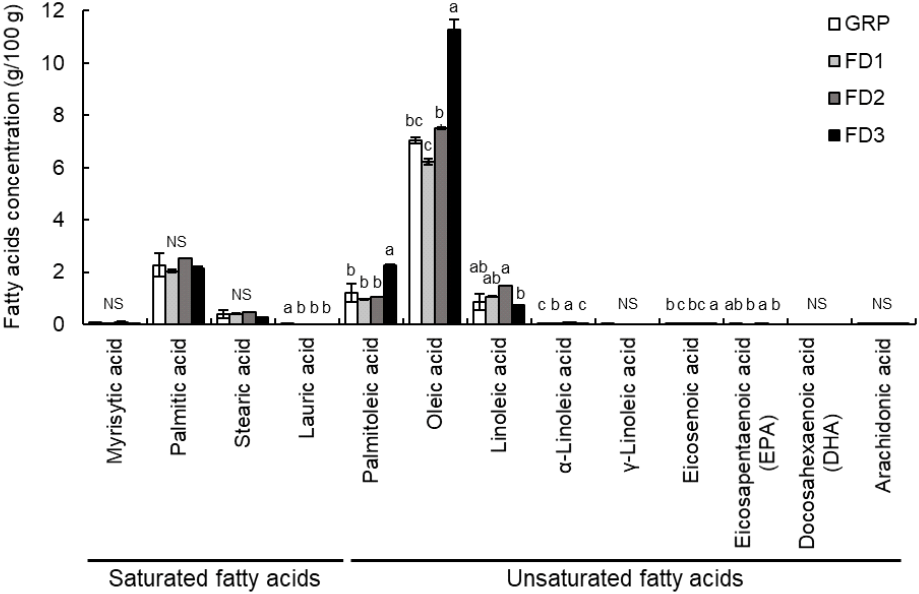
Among the remaining major fatty acids, palmitic acid (2.035-2.524 g/100 g), palmitoleic acid (0.960-2.246 g/100 g), and linoleic acid (0.726-1.486 g/100 g) were consistently abundant. Significant Tukey groupings were also observed for palmitoleic acid, linoleic acid, α-linolenic acid, eicosenoic acid, EPA, and lauric acid, whereas the remaining fatty acids were annotated as NS. Total quantified fatty acids were 11.959 g/100 g in GRP, 10.824 g/100 g in FD1, 13.237 g/100 g in FD2, and 16.819 g/100 g in FD3 larvae, with FD3 showing the highest total fatty acid value. When fatty acids were grouped by saturation class, the sum of saturated fatty acids (SFAs) ranged from 2.481 to 3.080 g/100 g, whereas the sum of unsaturated fatty acids (UFAs) ranged from 8.334 to 14.338 g/100 g, indicating that UFAs predominated across treatments (values calculated from Fig. 1).
Insects can exhibit diet-responsive fatty acid profiles, as dietary fatty acids and carbon sources can influence the relative abundance of specific fatty acids, including unsaturated fatty acids (Oonincx et al., 2015; Yoon et al., 2020). In P. brevitarsis larvae, variability in the distribution of major fatty acids—particularly oleic, palmitic, palmitoleic, and linoleic acids—has likewise been reported across rearing conditions and feeding substrates (Eom et al., 2024; Ham et al., 2021). Collectively, these results support the interpretation that formulated-diet pretreatment could serve as a controllable process variable to modulate selected fatty acids, as reflected by the Tukey letter groupings for oleic acid and other diet-responsive fatty acids.
Nevertheless, it cannot be determined conclusively whether the values in Fig. 1 primarily reflect physiological remodeling of lipid synthesis and/or storage during the pretreatment period, or whether they partly represent carryover effects from residual gut contents derived from the pretreatment diets. Although a 48-h starvation step was applied, the completeness of gut-content removal was not independently verified using quantitative indicators. Therefore, follow-up studies should integrate (1) the fatty acid composition of each diet, (2) quantitative measures of gut-emptying efficiency (e.g., frass output and residual gut-content mass), and (3) larval fatty acid composition to distinguish absorption and metabolic changes from residual-diet effects (Oonincx et al., 2015; Stanley‐Samuelson et al., 1988).
In the macromineral analysis (mg/100 g), Calcium (Ca) was significantly higher in all formulated-diet treatment groups than in GRP according to one-way ANOVA followed by Tukey’s HSD test. Ca increased from 243.7±6.2 mg/100 g in GRP to 694.1±45.3 mg/100 g in FD1, 644.1±47.1 mg/100 g in FD2, and 744.2±66.2 mg/100 g in FD3. Phosphorus (P) ranged from 993.5 to 1,191.5 mg/100 g in FD1-FD3 larvae and did not differ significantly from GRP larvae (1,015.0± 44.9 mg/100 g). Potassium (K) was significantly higher in FD1 larvae (3,407.6±81.4 mg/100 g) than in GRP larvae (3,089.5±79.9 mg/100 g) (p>0.05), whereas magnesium (Mg) did not differ significantly among treatments (388.0-428.5 mg/100 g). Regarding microminerals, zinc (Zn) was significantly higher in FD1 (11.5±0.4 mg/100 g) and FD2 (11.0±0.9 mg/100 g) than in GRP (8.8±0.0 mg/100 g), whereas FD3 was intermediate and did not differ significantly from either GRP or FD1-FD2 (Fig. 2B). Iron (Fe) was significantly higher in all FD groups than in GRP; among the FD groups, Fe was highest in FD1, lower in FD2, and intermediate in FD3.
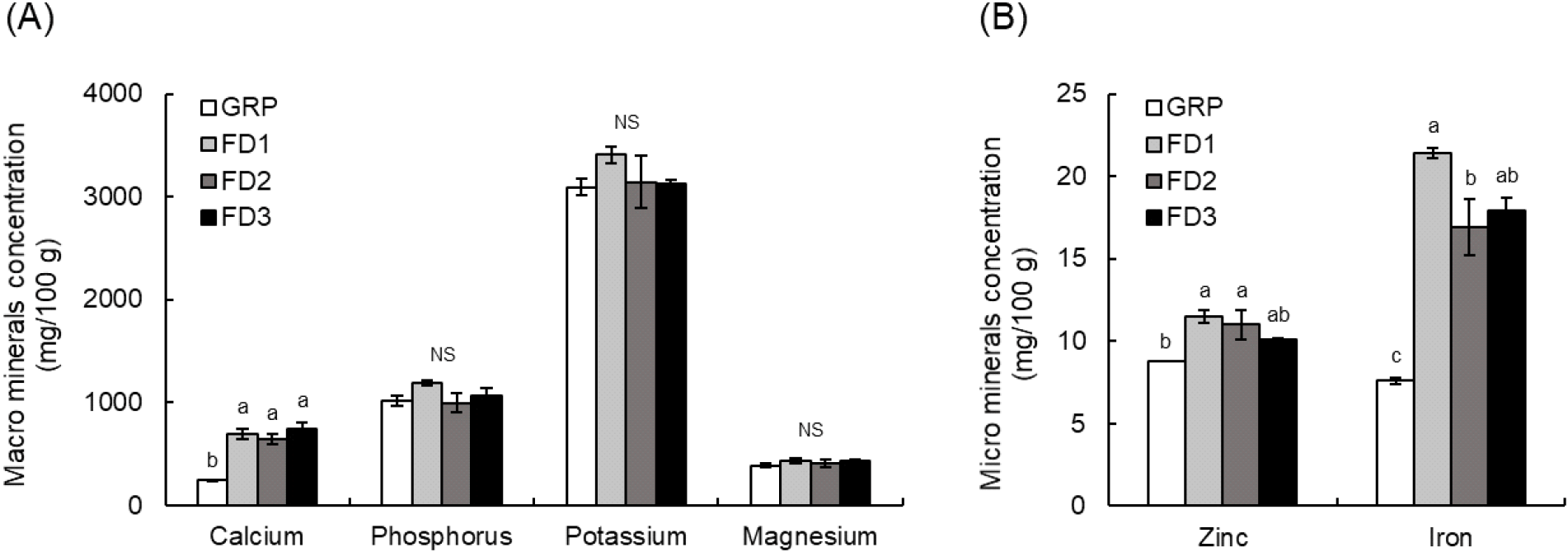
Overall, the FD pretreatments produced statistically supported increases in Ca and Fe (approximately 2.6-3.1-fold and 2.2-2.8-fold relative to GRP, respectively), whereas no significant treatment effects were detected for P, K, or Mg. Collectively, these results suggest that the short-term formulated diets used during the pretreatment step can alter—or potentially confound—the apparent mineral profile of P. brevitarsis larvae, particularly for Ca and Fe. Previous studies have reported that larval composition varies with feeding sources and rearing conditions (Ham et al., 2021; Yoon et al., 2020), and the present data further suggest that detectable shifts may occur even during brief gut-clearing treatments (Ham et al., 2021; Park et al., 2025a).
Notably, Ca was highest in FD3 despite FD3 having the lowest proportion of eggshell powder, indicating that the observed Ca increase cannot be explained solely by the eggshell inclusion rate. Several non-mutually exclusive factors may contribute, including baseline mineral contributions from other ingredients (e.g., wheat bran and brewer’s spent grain), variation in the extent of gut-content removal, and/or short-term redistribution following intake. To disentangle these effects, future studies should quantify mineral concentrations in individual ingredients (especially Ca, Fe, Zn, and K) and incorporate sampling strategies that more effectively separate larval tissue from residual gastrointestinal contents after pretreatment (Jankauskienė et al., 2024; Noyens et al., 2023).
Concentrations of Pb, Cd, and total As (mg/kg, dry weight) were low across all treatments (Table 3). Pb showed a significant Tukey grouping, with FD1-FD3 assigned to a higher group than GRP; however, the absolute difference was small (approximately 0.03 mg/kg). Cd and total As were annotated as NS, indicating no significant differences among treatment groups. Overall, the maximum observed concentrations were 0.069 mg/kg for Pb, 0.020 mg/kg for Cd, and 0.031 mg/kg for total As.
Abbreviations: GRP, larvae provided with the control diet (glutinous rice powder) during depuration (control group); FD1-FD3, larvae provided with formulated diets 1-3 during depuration (formulated-diet groups).
Because chemical hazards in insect-based foods—particularly toxic elements such as heavy metals—can be strongly influenced by the contamination profile of the rearing feed/substrate, monitoring is widely recognized as a key control point when feed ingredients or substrate sources change (EFSA Scientific Committee, 2015; van der Fels-Klerx et al., 2016). From a regulatory perspective, the Korean Food Code was revised under MFDS Notification No. 2021-54 to harmonize heavy-metal criteria across edible insect species and to regulate arsenic based on inorganic arsenic rather than total arsenic. Under these provisions for dried edible insects (dry-weight basis), the maximum levels are ≤0.1 mg/kg for Pb, Cd, and inorganic As, with a higher Pb limit (≤0.3 mg/kg) applied to P. brevitarsis larvae and Allomyrina dichotoma larvae (EFSA Scientific Committee, 2015; van der Fels-Klerx et al., 2018).
In Korea, the Food Code sets maximum levels for dried edible insects of 0.1 mg/kg for Pb (with a higher Pb limit of 0.3 mg/kg applied to P. brevitarsis larvae and Allomyrina dichotoma larvae), 0.1 mg/kg for Cd, and 0.1 mg/kg for As (MFDS, 2021). In this study, the maximum observed concentrations of Pb and Cd were 0.07 and 0.02 mg/kg, respectively, both well below these limits. Arsenic concentrations were also low, with total As not exceeding 0.03 mg/kg in any treatment and remaining below the 0.1 mg/kg regulatory threshold.
These results indicate that, despite the small statistically significant increase in Pb, the short-term formulated-diet gut-clearing pretreatment (FD1-FD3) did not introduce appreciable Pb, Cd, or As contamination under the present experimental conditions. Nevertheless, chemical hazards in edible insects depend strongly on the contamination profile of the rearing substrate and can exhibit element- and species-specific transfer and bioaccumulation (Malematja et al., 2023; Meyer et al., 2021; Truzzi et al., 2019; van der Fels-Klerx et al., 2016). Accordingly, for industrial application, changes in feed formulation or raw-material sourcing should be accompanied by routine screening of feed ingredients and lot-based monitoring of Pb, Cd, and As in both substrates and larvae.
The selected volatile compounds detected by the E-nose and their peak areas are summarized in Table 4, and the overall treatment-dependent variation is presented in the PCA score/loading plot in Fig. 3. In the loading plot, none of the selected discriminant volatiles were located in the first quadrant. Instead, 2,3-pentanedione and butanal clustered in the second quadrant; heptanal and butane-2,3-dione in the third quadrant; and anethole, 2-heptanone, and propyl propanoate in the fourth quadrant. All seven compounds showed DP values >0.90, indicating strong contributions to sample differentiation. PC1 and PC2 explained 99.992% and 0.007428% of the total variance, respectively, together accounting for 99.999428% of the overall variation. In the score plot, FD1 and FD3 were positioned close to each other, indicating similar overall volatile profiles. Conversely, the control group (GRP) was clearly separated from the formulated-diet treatments, and FD2 was also distinctly separated from the other groups and positioned closer to the fourth-quadrant marker compounds. Because odor-related attributes arise from complex mixtures of volatile compounds, this PCA pattern suggests that the depuration diets altered the overall odor-related characteristics of P. brevitarsis larvae rather than changing only a single volatile compound (Cai et al., 2024).
Volatile compounds were tentatively identified by electronic-nose analysis based on retention time and the instrument library.
Odor descriptors are library-based terms associated with the tentatively identified volatile compounds and do not represent sensory-panel data.
GRP, larvae provided with the control diet (glutinous rice powder) during the depuration step (control group); FD1-FD3, larvae provided with formulated diets 1-3 during the depuration step (formulated-diet groups).
Table 4 further supports the PCA-based separation. Butanal exhibited the largest peak area in GRP and was significantly lower in all formulated-diet groups, without a clear formulation-dependent trend. Butane-2,3-dione and heptanal were also highest in GRP and decreased in the formulated-diet groups, whereas 2,3-pentanedione was detected only in GRP. Conversely, propyl propanoate and 2-heptanone showed their highest peak areas in FD2, and anethole was detected exclusively in FD2. Taken together with the library-assigned odor descriptors in Table 4, these results indicate that GRP was more closely associated with butanal, 2,3-pentanedione, heptanal, and butane-2,3-dione, whereas FD2 was more closely associated with propyl propanoate, 2-heptanone, and anethole. This shift suggests that the conventional glutinous-rice-based control and the formulated depuration diets generated different volatile signatures, with FD2 showing the most distinctive treatment-specific pattern among the formulated-diet groups. The close placement of FD1 and FD3 in the score plot, despite differences in formulation, further suggests that their overall volatile patterns were more similar to each other than to GRP or FD2. Similar treatment-dependent changes in volatile composition have been reported for P. brevitarsis under different fermentation or rearing conditions, supporting the interpretation that the depuration diets used in the present study contributed to the observed differences in volatile profiles (Cha et al., 2024a; Cha et al., 2024b).
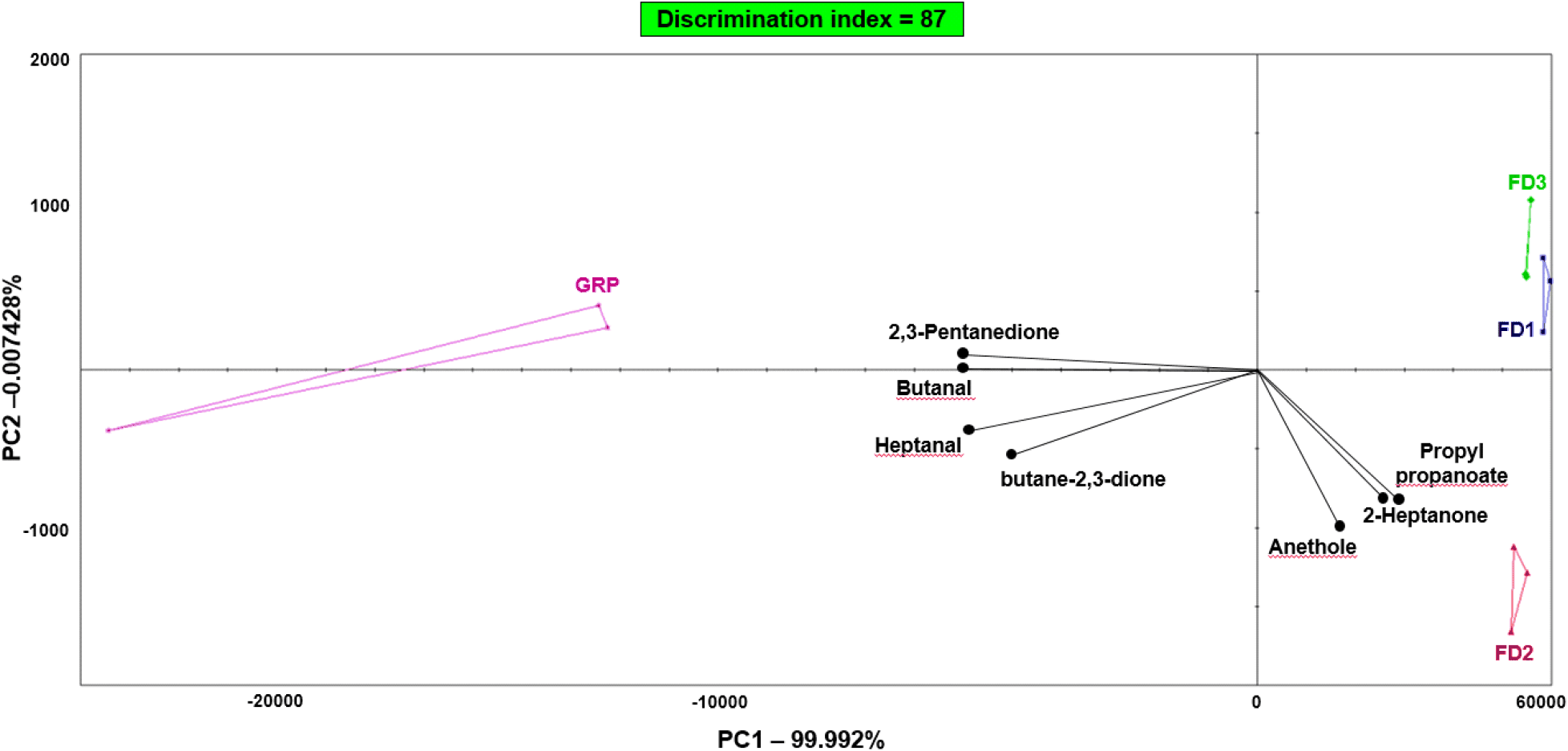
These findings are important because sensory quality remains a practical barrier to broader acceptance of insect-based foods, and odor profile is closely associated with consumer acceptance of P. brevitarsis larvae products (Cha et al., 2024a). However, the current E-nose approach is more suitable for rapid VOC fingerprinting and comparative pattern recognition than for definitive identification of key odorants. Accordingly, the odor descriptors in Table 4 should be interpreted as library-based tentative assignments rather than direct sensory-panel results (Cai et al., 2024).
In the present study, the formulated depuration diets did not simply reduce the abundance of selected GRP-associated compounds; rather, they shifted the dominant volatile markers toward a different profile. However, the odor descriptors listed in Table 4 are library-based terms for tentatively identified compounds and should not be interpreted as direct sensory-panel results. In addition, because the larvae were processed as whole-body materials and the analyzed samples were whole-larva powders obtained after 48 h of diet provision followed by 48 h of fasting, the observed differences may reflect both changes in larval tissue composition and differences in residual gut contents. Nevertheless, these data indicate that diet formulation during depuration can influence the volatile profile of P. brevitarsis larvae and may therefore serve as a practical pre-processing factor for modulating odor-related quality.
4. Conclusions
Formulated depuration diets altered the nutritional composition and volatile characteristics of P. brevitarsis larvae while maintaining Pb, Cd, and As contamination at negligible levels under the present conditions. Electronic-nose analysis further revealed clear diet-dependent separation of volatile profiles, with FD2 showing the most distinct pattern among the formulated-diet groups. These findings suggest that depuration diet formulation can serve as a practical pre-processing factor for modulating the quality of P. brevitarsis larvae, rather than functioning only as a gut-emptying step. However, because this study used a limited number of independent replicate containers, further validation with larger biological replication is required before broad industrial generalization.