
1. Introduction
Citrus fruits are well-known for their taste and nutritional benefits, but they are also packed with phytochemicals—health-promoting compounds that offer significant therapeutic potential. These secondary metabolites are also responsible for the fruit’s unique flavors and aromas. Among these, flavonoids are the most abundant group, with flavanones such as hesperidin and naringin being particularly characteristic of citrus (Fayek et al., 2019; Feng et al., 2018; Lee et al., 2022; Lin et al., 2023; Lv et al., 2015). These powerful phytochemicals exhibit a wide spectrum of health benefits, including potent antioxidant, anti-inflammatory, anti-cancer, anti-diabetic, and cardiovascular protective effects (Lv et al., 2015; Saini et al., 2022; Zou et al., 2016). Beyond flavonoids, citrus fruits contain other valuable metabolites such as limonoids (Saini et al., 2022), known for their anti-cancer properties and ability to stimulate detoxification enzymes (Kitagawa et al., 2021); carotenoids (Saini et al., 2022), which provide vibrant colors and act as antioxidants (Zou et al., 2016); and essential oils (González-Mas et al., 2019; Hee et al., 2006; Porat et al., 2016), rich in terpenes like limonene, offering antioxidant and antimicrobial attributes (Raspo et al., 2020). The synergistic action of these diverse metabolites underscores the holistic health advantages associated with regular citrus consumption, making them a crucial component of a healthy diet and a focus for nutraceutical research.
Inflammation is a fundamental immune response that acts as the body’s primary defense system. It is a complex process triggered to protect an organism from harmful stimuli, including pathogens, injured cells, and irritants (Malleshappa et al., 2018; Wang et al., 2018). Within this intricate process, macrophages play a pivotal role in orchestrating inflammatory responses against external aggressors (Pierce, 1990). When macrophages are activated by substances like lipopolysaccharide (LPS), they increase the production of pro-inflammatory cytokines, such as interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6). Concurrently, they synthesize inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), which in turn produce inflammatory mediators like nitric oxide (NO) (Ahmed, 2011; Miles and Calder, 2021; Wang et al., 2018). PGE2, another pro-inflammatory mediator, crucially suppresses the production of pro-inflammatory cytokines in macrophages (Yang et al., 2023). Given this, natural bioactive compounds, especially those prevalent in citrus fruits, are increasingly recognized as promising candidates for anti-inflammatory therapies. These phytochemicals act through modulation of inflammatory signaling pathways, making them attractive for developing novel interventions. Evidence from multiple studies supports their ability to improve skin health by reducing inflammation and enhancing tissue repair. Among these, polymethoxyflavonoids (PMFs) and their hydroxylated derivatives (HPMFs) have received particular attention for their anti-inflammatory potential (Fontana et al., 2023; Wang et al., 2018), other flavonoids (Azuma et al., 2020; Benavente-García and Castillo, 2008), essential oils (Amorim et al., 2016; Yang et al., 2023), and vitamin C (Miles and Calder, 2021). Furthermore, various extracts derived from citrus fruits have also demonstrated their potential as sources of anti-inflammatory agents (Lee et al., 2013; Malleshappa et al., 2018).
Sarahyang, a relatively new citrus cultivar, is originated from the nucellar tissue of the popular citrus hybrid Setoka (Woo et al., 2020). Developed by the Citrus Research Center of the National Institute of Horticultural and Herbal Science, Rural Development Administration (RDA) in Korea, this cultivar was created in 2001 and officially introduced around 2017. Sarahyang is recognized for several desirable traits that aim to improve upon existing citrus varieties, including an average fruit weight of about 200 g, with both peel and flesh displaying an appealing orange color when mature. It stands out for its strong sweetness, boasting a reported °Brix level of 14.0 °Brix at maturity, which is notably 1.0-1.5 °Brix higher than its maternal parent, Setoka. Concurrently, its acid content at maturity is around 1.0%, a level 0.5-1.0% lower than that of Setoka. This combination of high sugar and relatively low acid contributes significantly to Sarahyang’s appealing taste. Furthermore, it matures earlier than Setoka, typically in mid to late February—approximately one-month sooner—and is characterized by abundant juice and excellent texture. Its peel thickness is comparable to that of Satsuma mandarin, and it is generally easy to peel. While its tree vigor is similar to Setoka, Sarahyang tends to have a high fruit-setting rate, which can necessitate careful management to prevent over-fruiting (Park et al., 2025).
At the Citrus Research Center, our continuous breeding program is dedicated to developing premium homegrown citrus cultivars that excel in taste, aroma, and seedlessness, while also demonstrating strong commercial viability. Among the new cultivars developed through this program, Sarahyang represents one of our latest advancements. To thoroughly understand Sarahyang and its potential benefits, we conducted a comprehensive metabolite and bioactivity study. This research involved evaluating the metabolite dynamics of the fruit over six months, specifically from October to March, to track changes in its chemical composition during ripening and investigated the anti-inflammatory properties of Sarahyang extracts (SE) by testing their impact on inflammation in RAW 264.7 mouse macrophage cells. This dual approach helps us understand both the superior quality and potential health benefits of this promising new domestic cultivar.
2. Materials and methods
The study on the new mandarin cultivar, Sarahyang, was conducted at the National Institute of Horticultural and Herbal Science Citrus Research Center in Seogwipo-si, Jeju-do, with samples collected monthly from October 2022 to March 2023. Different preparation procedures were applied depending on the intended analysis. The pulp was juiced to determine free sugar and organic acid composition. For assessments of flavonoid content and anti-inflammatory properties, both pulp and peel were separated, dried at 55°C, and extracted using 70% ethanol at a solvent-to-sample ratio of 20:1 (v/w). The extracts underwent sonication for 1 h at 37°C, followed by centrifugation, filtration, and concentration at 45°C under reduced pressure. The concentrated solutions were subsequently freeze-dried to obtain stable extracts for further experiments. Specifically, peels from fully ripened Sarahyang fruits were used to investigate anti-inflammatory activity.
Fruit weight was measured with an electronic balance, while length and width were determined using a Vernier caliper. Total soluble solids (TSS) content, measured in °Brix, was obtained from the supernatant of fresh flesh juice using a digital refractometer after centrifugation at 10,000 ×g for 15 min at 4°C. Titratable acidity (TA) was calculated as citric acid equivalent after titrating a diluted sample of the supernatant (2 mL in 8 mL distilled water) with 0.1 N NaOH to a pH of 8.1.
Free sugars (sucrose, glucose, and fructose) and organic acids (oxalic, malic, and citric acids) were quantified using high-performance liquid chromatography (HPLC). For analysis, the juice was diluted tenfold and passed through a 0.45 μm membrane filter prior to injection. Sugar separation was carried out on a ZORBAX NH2 C18 column (4.6×250 mm, 5 μm; Agilent) using an isocratic elution and mobile phase of acetonitrile and water (75:25, v/v), at a flow rate of 1 mL/min and column temperature of 40°C. Detection was performed with a refractive index detector (RID-20A, Shimadzu), and quantification was achieved through external calibration with authentic sugar standards. Organic acids were analyzed on the same instrument, employing a Shim-Pak GIS C18 column (4.6×250 mm, 5 μm; Shimadzu). The mobile phase consisted of solvent A (10 mM sodium phosphate buffer, pH 2.6) and solvent B (acetonitrile), with gradient elution programmed as follows: 100% A for 10 min, transition to 10% A/90% B over 7 min, then 100% B for 13 min. The flow rate was 1 mL/min, the column temperature was held at 30°C, and detection was achieved at 226 nm using a UV-visible detector. Concentrations of organic acids were determined from calibration curves constructed with standard compounds. Solvents and chemicals such as acetonitrile, sodium phosphate buffer, standards of sugar (sucrose, glucose, and fructose) and organic acids (oxalic, malic, and citric acids) used in HPLC analyses were HPLC grade and purchased from Sigma-Aldrich (St. Louis, MO, USA).
For flavonoid profiling, the freeze-dried extracts were reconstituted to the required concentration, filtered through a 0.22 μm PVDF membrane, and analyzed using a Waters e2695 HPLC system equipped with a UV-Vis detector (Waters 2489) and a YMC-Triart C18 column (250×4.6 mm, 5 μm). Chromatographic separation was performed at a flow rate of 1 mL/min under isocratic conditions, with solvent composition adjusted according to the flavonoid group: glycosylated flavonoids (rutin, narirutin, hesperidin) were separated with acetonitrile and 20 mM phosphoric acid (2:8, v/v) and polymethoxylated flavonoids (nobiletin, tangeretin) with acetonitrile and 20 mM phosphoric acid (6:4, v/v). The column temperature was maintained at 45°C, and detection was carried out at 280 nm. Flavonoids were identified by comparing retention times and UV spectra with authentic standards. Quantification was performed using external calibration curves prepared for each standard across the concentration range of 15.625-1,000 μg/mL. Standards of rutin, narirutin, hesperidin, nobiletin, and tangeretin were HPLC grade and purchased from Sigma-Aldrich. The final value were reported in mg/100g of the dried extracts (DE).
RAW 264.7 murine macrophages were obtained from the Korea Cell Line Bank (KCLB, Seoul, Korea) and cultured in Dulbecco’s Modified Eagle Medium (DMEM; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Merck Millipore, Burlington, MA, USA) and 1% penicillin-streptomycin (Thermo Fisher Scientific). Cells were maintained at 37°C in a humidified incubator with 5% CO2 and 95% air. The medium was replaced every two days to sustain healthy cell growth.
Cell viability was determined using a modified MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay, following a previously reported method with slight modifications (Yoon et al., 2009). RAW 264.7 cells (1×105 cells/well) were seeded into 24-well plates and treated with SE at concentrations of 100, 200, 400, and 800 μg/mL in the presence of 1 μg/mL LPS for 24 h. After treatment, the medium was replaced with 400 μL of MTT solution (0.2 mg/mL) and incubated for 3 h at 37°C. The supernatant was then discarded, and the resulting formazan crystals were dissolved in 800 μL of dimethyl sulfoxide (DMSO). Absorbance was measured at 570 nm using a SpectraMaxⓇ M3 Multi-Mode Microplate Reader (Molecular Devices, Sunnyvale, CA, USA). Cell viability was calculated relative to untreated control cells and expressed as mean±standard deviation (SD) from three independent experiments.
Cells were seeded at a density of 1.5×105 cells per well in a 24-well plate and treated with SE at concentrations of 25, 50, 100, 200, and 400 μg/mL. Simultaneously, cells were stimulated with 1 μg/mL of LPS and incubated for 24 h at 37°C. After incubation, the culture media were collected from each well to assess nitrite levels using the Griess reagent, composed of 1% sulfanilamide, 0.1% N-(1-naphthyl) ethylenediamine, and 2.5% phosphoric acid. Equal volumes of culture medium (100 μL) and Griess reagent (100 μL) were mixed and incubated at room temperature (18-25°C) for 15 min. Absorbance was measured at 540 nm using a SpectraMaxⓇ M3 Multi-Mode Microplate Reader (Molecular Devices). Results are presented as the mean±SD from three independent experiments conducted in triplicate.
RAW 264.7 cells were plated at a density of 1.5×105 cells per well in 500 μL of culture medium using a 24-well plate and allowed to pre-incubate for 24 h at 37°C in a humidified incubator with 5% CO2. Following this initial incubation, cells were exposed to increasing concentrations of SE (50, 100, 200, and 400 μg/mL). After SE treatment, LPS was added at a final concentration of 1 μg/mL, and cells were incubated for an additional 24 h. Levels of PGE2, IL-6, and TNF-α in the collected supernatants were measured using commercially available ELISA kits, following the protocols provided by the manufacturer.
Nuclear and cytoplasmic extraction was performed using a commercial kit on RAW 264.7 cells. The cells (6.0×105 cells/dish) were cultured for 24 h in 60 mm dishes before being treated with SE and LPS (1 μg/mL) to induce protein expression. The nuclear extract was then isolated following the manufacturer’s protocol.
Western blot analysis was performed with slight modifications according to previously reported method (Han and Hyun, 2023). In brief, RAW 264.7 cells were seeded at a density of 6.0×105 cells per 60 mm culture dish in 3 mL of medium and pre-incubated for 1 h at 37°C in a 5% CO2 atmosphere. The cells were then treated with SE at concentrations of 100, 200, and 400 μg/mL, along with LPS (1 μg/mL), according to the required expression time of the target proteins. Following treatment, cells were lysed, and total protein concentrations were normalized to 30 μg/mL. Protein samples were denatured using Laemmli buffer, resolved by SDS-PAGE, and subsequently transferred onto PVDF membranes. Membranes were blocked in 5% skim milk and incubated overnight at 4°C with primary antibodies diluted 1:1000. After washing, membranes were incubated with HRP-conjugated secondary antibodies (1:1000) for 2 h at room temperature. Protein bands were detected using an enhanced chemiluminescence (ECL) system and visualized with the ChemiDoc imaging system (Vilber Lourmat, France). The following primary antibodies were used: p-ERK (9101S), ERK (9102S), p-p38 (9211S), p38 (9212S), p-JNK (9251S), p-IκBα (9246S), IκBα (4812S), p65 (4764S), and β-actin (4967S), all from Cell Signaling Technology (Danvers, MA, USA). Anti-iNOS (2982S) was sourced from Merck Millipore (Burlington, MA, USA), and anti-COX-2 was obtained from BD Biosciences (Franklin Lakes, NJ, USA). Secondary antibodies (anti-mouse and anti-rabbit) were also purchased from Cell Signaling Technology. ImageJ program was used to quantify the intensities of western blot bands, which is displayed in the graph.
All experimental data are presented as mean±SD from triplicate experiments. For physicochemical analysis, one-way ANOVA followed by Tukey’s honest significant difference (HSD) test was performed using R-Software (R4.5.1). For anti-inflammatory tests, statistical analysis was conducted using Student’s t-test. Significance probability were denoted as follows: p<0.05 (*), p<0.01 (**), and p<0.001 (***) when compared to the LPS-treated group, and p<0.001 (#) when compared to the untreated control group.
3. Results and discussion
The maturation of the citrus fruit, observed from October to March, exhibited distinct and interrelated trends in its quality parameters (Table 1). TSS, an indicator of sweetness, generally increased throughout the period, rising from 7.9 °Brix in October to a peak of 14.4 °Brix by late March, despite a slight dip in January. Conversely, TA consistently decreased, starting at 3.53% in October and falling to 0.93% by late March, demonstrating a very strong negative correlation with TSS (-0.941). These observations regarding TA and TSS dynamics are consistent with similar studies involving sweet fruits, including pomegranate accessions (Shwartz et al., 2009) and oranges (Grilo et al., 2017). Conversely, the findings show a discrepancy with literature on acidic citrus varieties (lime, lemon, and orange), which report an initial increase in TA during early fruit development, followed by constant levels in acidless varieties (Albertini et al., 2006). Concurrently, the fruit’s physical dimensions—weight, length, and height—generally increased from October to February (peaking at 239.9 g, 80.9 mm, and 62.9 mm respectively), before experiencing a notable, though temporary, decrease in early March and then rebounding by late March. The very strong positive correlations among weight, length, and height confirm the proportional growth of the fruit in all dimensions.
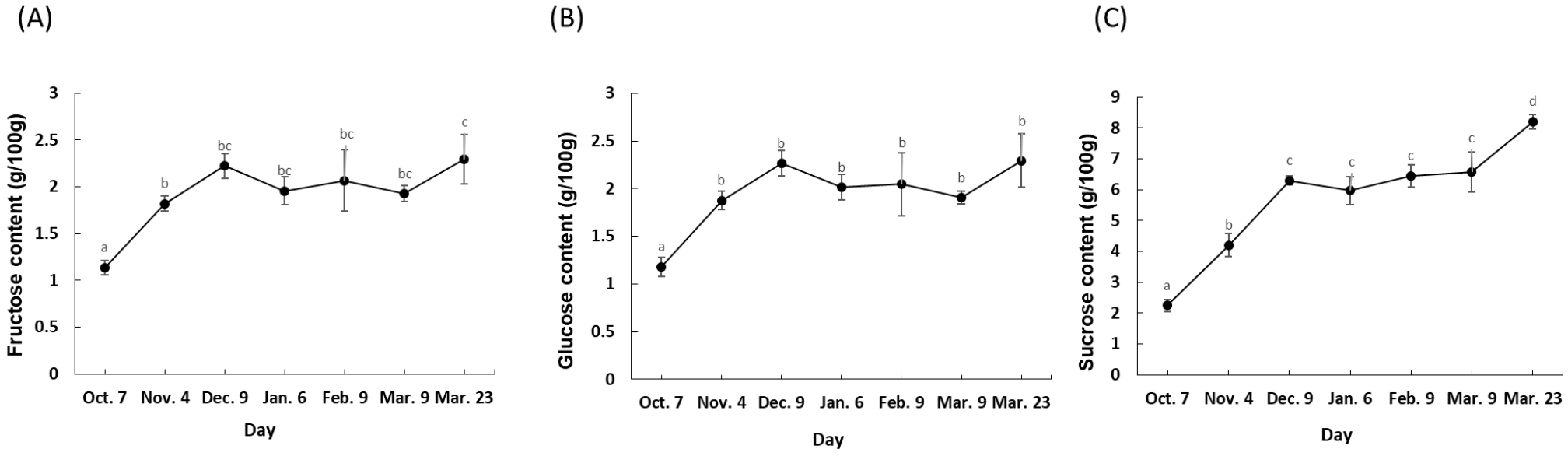
The analysis of free sugar concentrations in the Sarahyang fruit juice over its maturation period reveals clear trends and strong interrelationships among individual sugars (Fig. 1). Fructose and glucose exhibited highly similar patterns. Both generally increased from October (fructose: 1.1 g/100 g, glucose: 1.2 g/100 g) to a peak in December (fructose: 2.2 g/100 g, glucose: 2.3 g/100 g). Following a slight dip in January, their levels remained relatively stable through February and early March, before rising again to their highest points by late March (fructose: 2.3 g/100 g, glucose: 2.3 g/100 g). Sucrose, however, showed a more consistent and pronounced increasing trend throughout the entire measurement period, starting at 2.2 g/100 g in October and reaching its highest concentration of 8.2 g/100 g by late March, despite a minor decrease in January. This indicates that sucrose accumulates significantly as the fruit matures, contributing most to the increasing sweetness. As free sugars (specifically glucose, fructose, and sucrose) are the primary constituents of the TSS measurement, the observed increase is a consequence of the fruit’s ripening profile. This trend aligns with findings reported for other sweet fruit varieties, including orange (Albertini et al., 2006), mandarins (Bermejo and Cano, 2012), and pomegranate (Shwartz et al., 2009). In sharp contrast, this pattern diverges from the developmental profiles documented for highly acidic citrus cultivars (e.g., lime and lemon), where inconsistent sugar accumulation trends have been reported (Albertini et al., 2006). The correlation analysis further highlights these relationships: fructose and glucose are extremely strongly positively correlated (0.996), indicating they accumulate and fluctuate almost in unison. Sucrose also shows strong positive correlations with both fructose (0.928) and glucose (0.902).
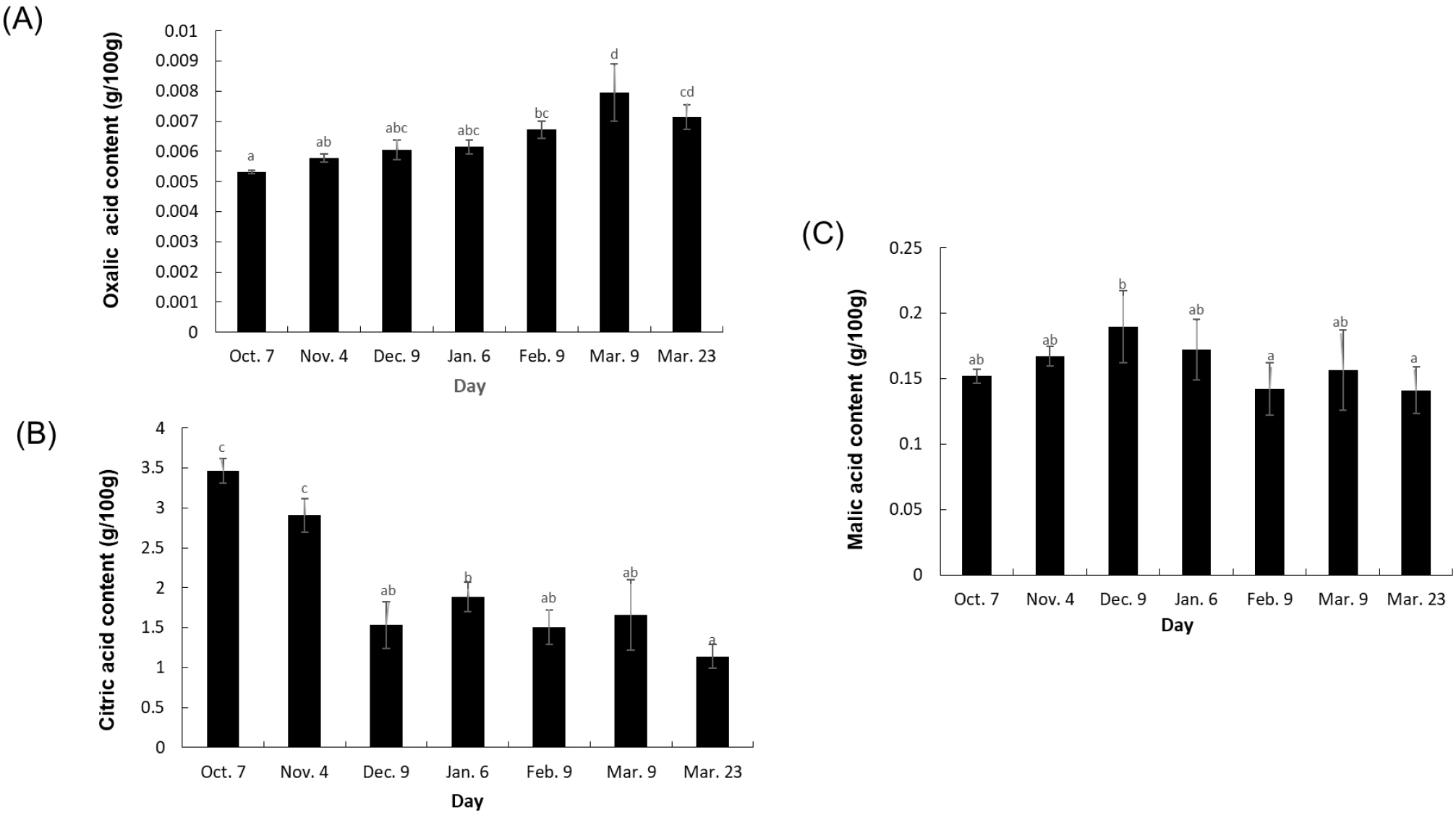
The analysis of organic acid concentrations in the Sarahyang fruit juice over time revealed distinct patterns for each acid (Fig. 2). Citric acid, the most abundant, displayed a consistent and significant decreasing trend from 3.5 g/100 g in October to its lowest point of 1.1 g/100 g by late March, a key indicator of ripening. The accumulation dynamics of organic acids appear to be highly variety-dependent. The observed decrease in citric acid levels is consistent with the overall decline in TA, given citric acid’s significant contribution to TA. This result concurs with findings from a previous study on pomegranate (Shwartz et al., 2009). Conversely, this finding contrasts with the developmental profile of acidic citrus varieties (lime, lemon, and orange), where citric acid levels are often reported to increase in the early stages of development before decreasing later (Albertini et al., 2006). On the other hand, oxalic acid generally showed an increasing trend, rising from 0.0053 g/100 g to 0.0079 g/100 g by early March. This increasing pattern aligns with trends previously reported in lime, lemon, and orange varieties (Albertini et al., 2006; Bermejo and Cano, 2012). Meanwhile, malic acid concentrations exhibited a more fluctuating pattern, peaking in December (0.1897 g/100 g) before generally declining through February and late March (0.1411 g/100 g). While the levels of malic acid remained relatively constant during the development of lime, lemon, and orange varieties (Albertini et al., 2006), an increasing trend was documented in certain pomegranate accessions (Shwartz et al., 2009). Regarding relationships, a moderately strong negative correlation (-0.726) was observed between citric acid and oxalic acid, suggesting that as citric acid diminishes during ripening, oxalic acid tends to accumulate. In contrast, the correlation between citric acid and malic acid was very weak and positive (0.062), indicating little linear relationship between their changes. A weak negative correlation (-0.364) existed between oxalic and malic acids, though this relationship was not prominent. Collectively, these differential patterns in organic acid metabolism, especially the pronounced decrease in citric acid, are crucial contributors to the evolving taste profile of the fruit throughout its maturation.
The analysis of major flavonoids concentrations in the Sarahyang fruit’s pulp and peel from October to March reveals distinct trends, significant differences between tissues, and strong positive interrelationships among the compounds (Quantitative values are presented in Table 2). Across the measurement period, concentrations of rutin, narirutin, and hesperidin generally declined in the pulp. This decline was particularly significant for narirutin, which dropped from an initial 1,629.5 mg/100 g to 555.7 mg/100 g by late March. Similarly, all measured flavonoids in the peel exhibited a clear decreasing trend, exemplified by the steep reduction in narirutin concentrations from 13,810.0 mg/100 g to 4,391.0 mg/100 g. Comparative studies on flavonoid accumulation during citrus ripening have yielded inconsistent results, although the general decreasing trend observed in Sarahyang aligns with several reports. Specifically, high levels of flavonoids, including hesperidin, rutin, and nobiletin, have been documented at the early stages of development for numerous late-maturing citrus varieties, followed by a rapid decrease in both the peel (Kim et al., 2002) and the juice (Kim et al., 2009) Furthermore, the consistent decrease of narirutin with maturity has also been reported in grapefruit (Chaudhary et al., 2016). However, another report indicated divergent accumulation patterns. For example, the peel of the ‘Valencia’ orange showed narirutin decreasing while hesperidin, nobiletin, and tangeretin increased with maturation (Grilo et al., 2017). Conversely, one study found that levels of rutin and nobiletin increased early and then decreased later, which was the opposite tendency observed for narirutin (Zhu et al., 2020). This overall changes in flavonoid content with maturity is often attributable to the down and up-regulation of genes related to their synthesis in different stage of the fruits (Chaudhary et al., 2016). Our analysis of the flavonoids revealed that the peel consistently contained significantly higher concentrations of all detected flavonoids compared to the pulp, with average concentrations for narirutin being over eight times higher in the peel (7,173.5 mg/100 g) than in the pulp (855.3 mg/100 g). Furthermore, nobiletin and tangeretin were exclusively found in the peel, with average concentrations of 958.6 mg/100 g and 56.0 mg/100 g, respectively, highlighting this tissue as the primary reservoir. Previous reports also showed that the peel is consistently found to be the most concentrated source of these bioactive compounds (Jing et al., 2015; Lee et al., 2022; Malleshappa et al., 2018). Within both the pulp and peel, the detected flavonoids exhibited extremely strong positive correlations with each other (e.g., rutin and narirutin at 0.982 in the pulp, and nobiletin and tangeretin at 0.999 in the peel), suggesting highly synchronized metabolic patterns.
Citrus fruit is an important horticultural crop that possess various natural metabolites which potentially provides nutraceutical and cosmetological benefits (Leong et al., 2024; Lv et al., 2015; Saini et al., 2022). The peel is the exclusive source of nobiletin and tangeretin, both of which are polymethoxyflavones often used in cosmetic and medicinal applications (Fontana et al., 2023; Leong et al., 2024; Wang et al., 2018). Citrus peel essential oils and extracts from fruits like lemons and oranges exhibit potent anti-inflammatory effects in both in vitro and in vivo models (Desmiaty et al., 2024; Lee et al., 2013; Malleshappa et al., 2018; Yang et al., 2023). Prior studies have repeatedly demonstrated that citrus fruits and their extracts have anti-inflammatory properties, which could be likely due to their high content of flavonoids and limonoids, compounds that have consistently shown positive results in anti-inflammatory tests (Amorim et al., 2016; Azuma et al., 2020; Fontana et al., 2023; Yang et al., 2023). The five compounds identified in Sarahyang peel—rutin, narirutin, hesperidin, nobiletin, and tangeretin—are all associated with established anti-inflammatory properties (Alsharairi, 2025; Desmiaty et al., 2024; Ingegneri et al., 2024). For example, nobiletin demonstrated potent inhibitory activity, blocking the LPS-induced secretion of key proinflammatory mediators (NO, TNF-α, IL-1b, and IL-6) by greater than 50% at a concentration of 100 μM. Furthermore, while hesperidin, nobiletin, and tangeretin individually exhibited only a mild anti-inflammatory effect, their synergistic combination resulted in a significant suppression of LPS-induced proinflammatory cytokine expression(Ho and Kuo, 2014). Narirutin has been reported to modulate multiple molecular targets, including the regulation of various inflammatory cytokines (Pandey et al., 2024). Complementary to these findings, rutin was shown to exert strong efficacy by potently inhibiting HMGB1 release and subsequently down-regulating the resulting HMGB1-dependent inflammatory responses in human endothelial cells (Yoo et al., 2014). Clinical studies provide further support, as daily consumption of orange juice has been shown to reduce systemic markers of inflammation, such as C-reactive protein (CRP), confirming the direct role of these fruit components in reducing inflammation (Coelho et al., 2013; Rocha et al., 2017). Citrus extract could exert its anti-inflammatory effects on lipopolysaccharide (LPS)-stimulated RAW 264.7 cells primarily by targeting and inhibiting key signaling pathways and downstream inflammatory mediators. This process is a coordinated cascade that ultimately prevents the transcription and production of pro-inflammatory substances.
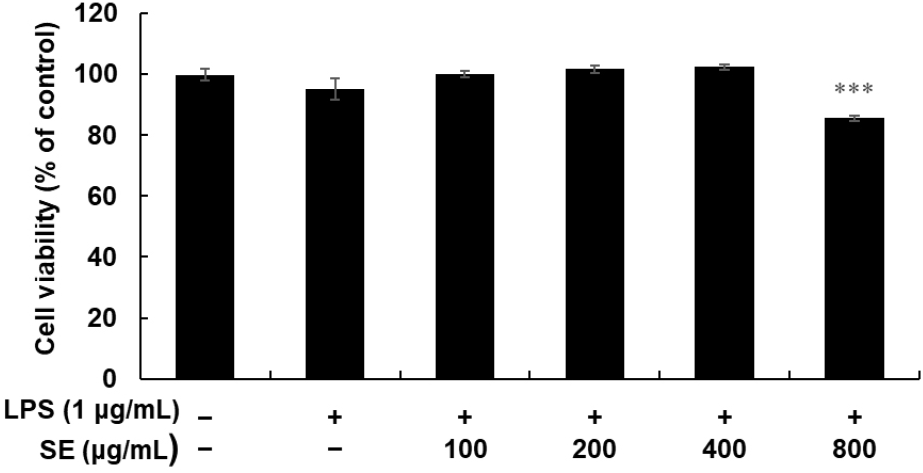
Ensuring no detrimental effects on cellular metabolism is crucial to study the biological activity of plant extracts. To investigate the appropriate concentration of SE for subsequent experiments, its cytotoxic effect on RAW 264.7 cells was determined using an MTT assay (Fig. 3). The viability test involved incubating cells with SE concentrations ranging from 100 to 800 μg/mL for 24 h, in the presence of 1 μg/mL LPS. Our results indicated that 800 μg/mL of SE significantly affected the viability of RAW 264.7 cells compared to untreated cells. However, concentrations lower than 400 μg/mL showed no cytotoxicity. Consequently, concentrations below 400 μg/mL were used to evaluate the anti-inflammatory effects of SE in RAW 264.7 cells.
Changes in the physiology of macrophages due to inflammation is mainly the result of activation of NF-κB and MAPK signaling pathways. The transcription factor NF-κB plays a key role in the immune response to LPS-induced inflammation by regulating the transcription of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 and enzymes like iNOS and COX-2 (Liu et al., 2017). This anti-inflammatory action is driven by a coordinated inhibition of crucial signaling pathways. SE was shown to suppress the expression of the enzymes iNOS and COX-2, which are responsible for producing NO and PGE2, respectively. SE also significantly inhibited LPS-induced degradation of IκB-α and the NF-κB p65 translocation from cytosol to nucleus. The underlying mechanism involves the regulation of the NF-κB signaling pathway, where SE prevents the activation of the p65 subunit, thereby blocking the transcription of inflammatory genes. The effect of SE on cell viability, production of NO, PGE2, pro-inflammatory cytokines, enzymes (iNOS and COX-2), and MAPKs are detailed in the following sections.
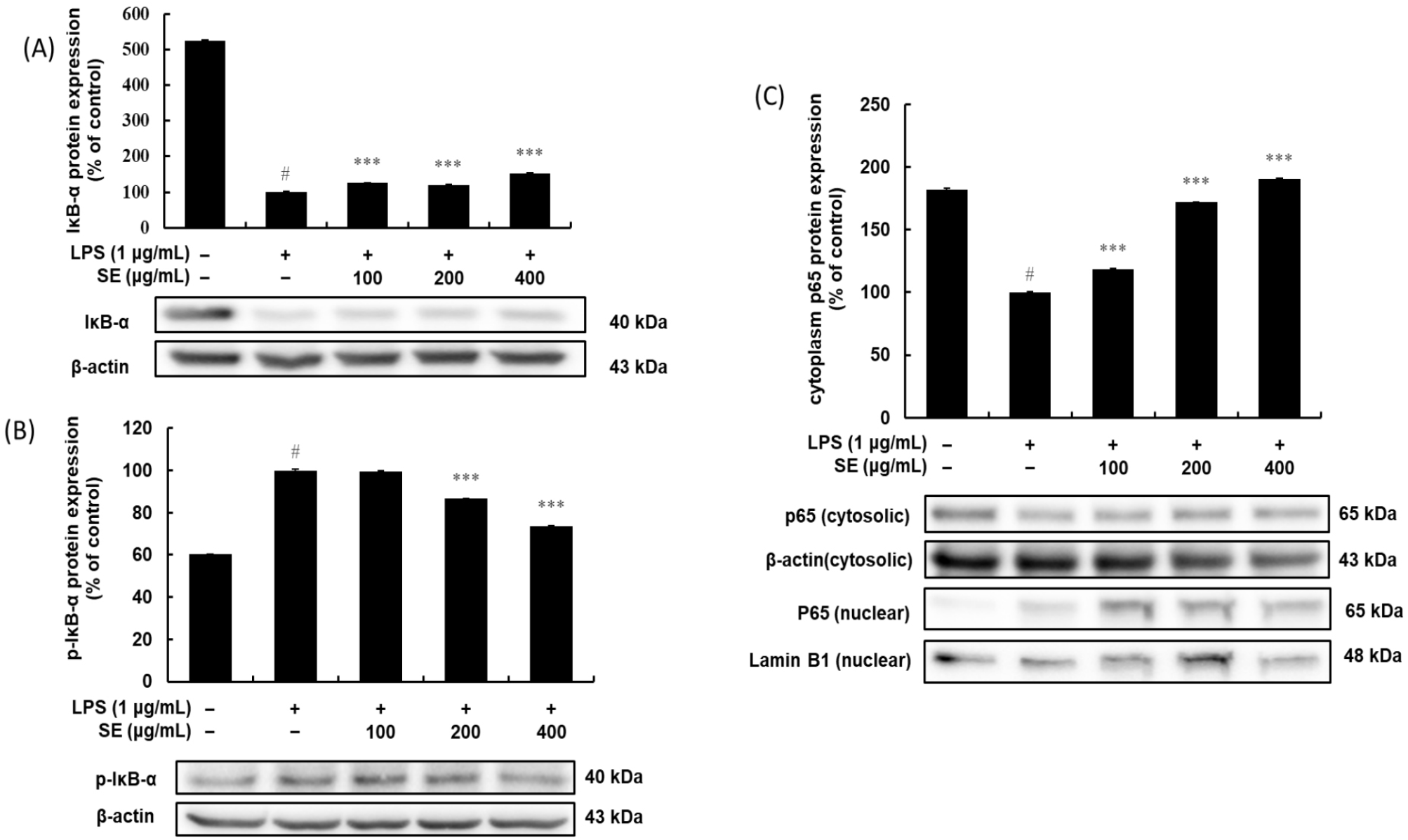
The results shown in Figs. 4A-4C indicate that SE effectively inhibits the NF-κB signaling pathway, which is a key regulator of inflammation. The pathway’s inhibition is clearly shown through the regulation of IκB-α and the p65 subunit. In unstimulated cells, a high level of IκB-α is present, which keeps the p65 subunit inactive in the cytoplasm (Figs. 4A and 4C). However, upon LPS stimulation, IκB-α undergoes rapid phosphorylation (Fig. 4B) and subsequent degradation (Fig. 4A), which frees p65 to translocate into the nucleus, as evidenced by the significant decrease in its cytoplasmic levels (Fig. 4C). SE treatment reverses this entire cascade in a dose-dependent manner, suppressing the LPS-induced phosphorylation of IκB-α (Fig. 4B) and, as a result, preventing its degradation (Fig. 4A). By preserving IκB-α, SE successfully sequesters the p65 subunit in the cytoplasm, blocking its translocation to the nucleus (Fig. 4C) and its ability to activate pro-inflammatory genes.
The flavonoids exert anti-inflammatory effects by modulating signaling cascades, most notably the NF-κB pathway, reducing inflammatory enzymes, and acting as antioxidants (Fontana et al., 2023; Miles and Calder, 2021). Limonoids, another class of compounds found particularly in seeds and peels of citrus (González-Mas et al., 2019), are well-known compounds exhibiting anti-inflammatory properties, often by inhibiting the NF-κB pathway and reducing pro-inflammatory cytokine expression (Amorim et al., 2016; Yang et al., 2023). The NF-κB pathway is a central regulator of inflammation and a primary target for many plant-derived compounds. In its inactive state, the NF-κB complex, which includes the p65 subunit, resides in the cytoplasm bound to its inhibitor protein, IκB-α. LPS stimulation triggers a phosphorylation cascade that leads to the phosphorylation and subsequent degradation of IκB-α. This degradation releases p65, allowing it to translocate into the nucleus. Once in the nucleus, p65 binds to DNA and initiates the transcription of numerous pro-inflammatory genes. The anti-inflammatory agents often work by inhibiting the phosphorylation of IκB-α, thereby preventing its degradation and keeping p65 sequestered in the cytoplasm. This blocks the entire downstream inflammatory response, suppressing the expression of the enzymes iNOS and COX-2 and cytokines such as TNF-α and IL-6. The suppression of iNOS and COX-2 would in turn limit the synthesis of inflammatory mediators (PGE2 and NO) that are involved in pain and fever (Liu et al., 2017; Moynagh, 2005).
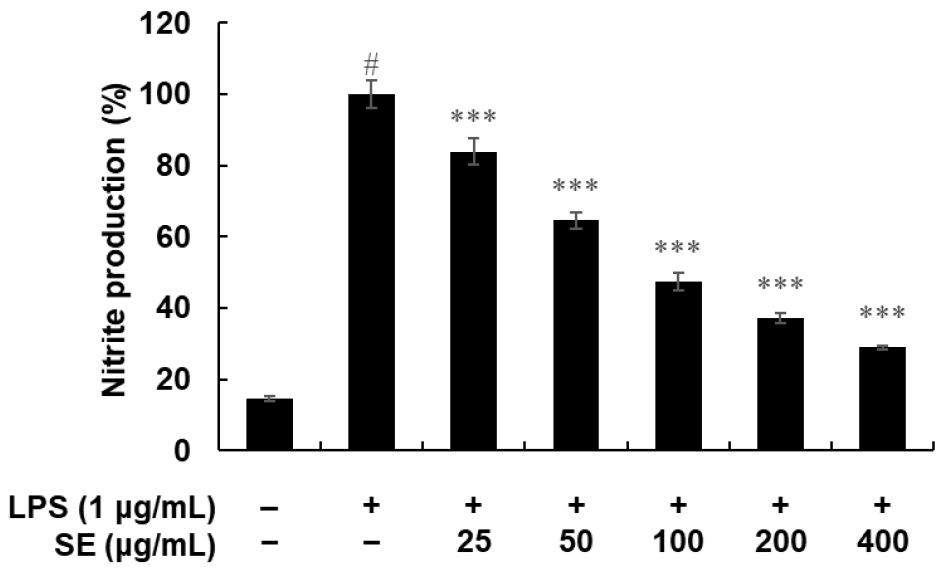
The amount of nitrite was determined using the Griess reaction (Fig. 5). In unstimulated RAW 264.7 cells, NO production was minimal, below 20%. In contrast, LPS treatment alone led to a marked increase in NO production. SE notably reduced this LPS-induced NO production in a dose-dependent manner. Similarly, PGE2 levels in the medium, which were significantly elevated upon LPS treatment, were also inhibited by SE in a dose-dependent fashion (Fig. 6A).
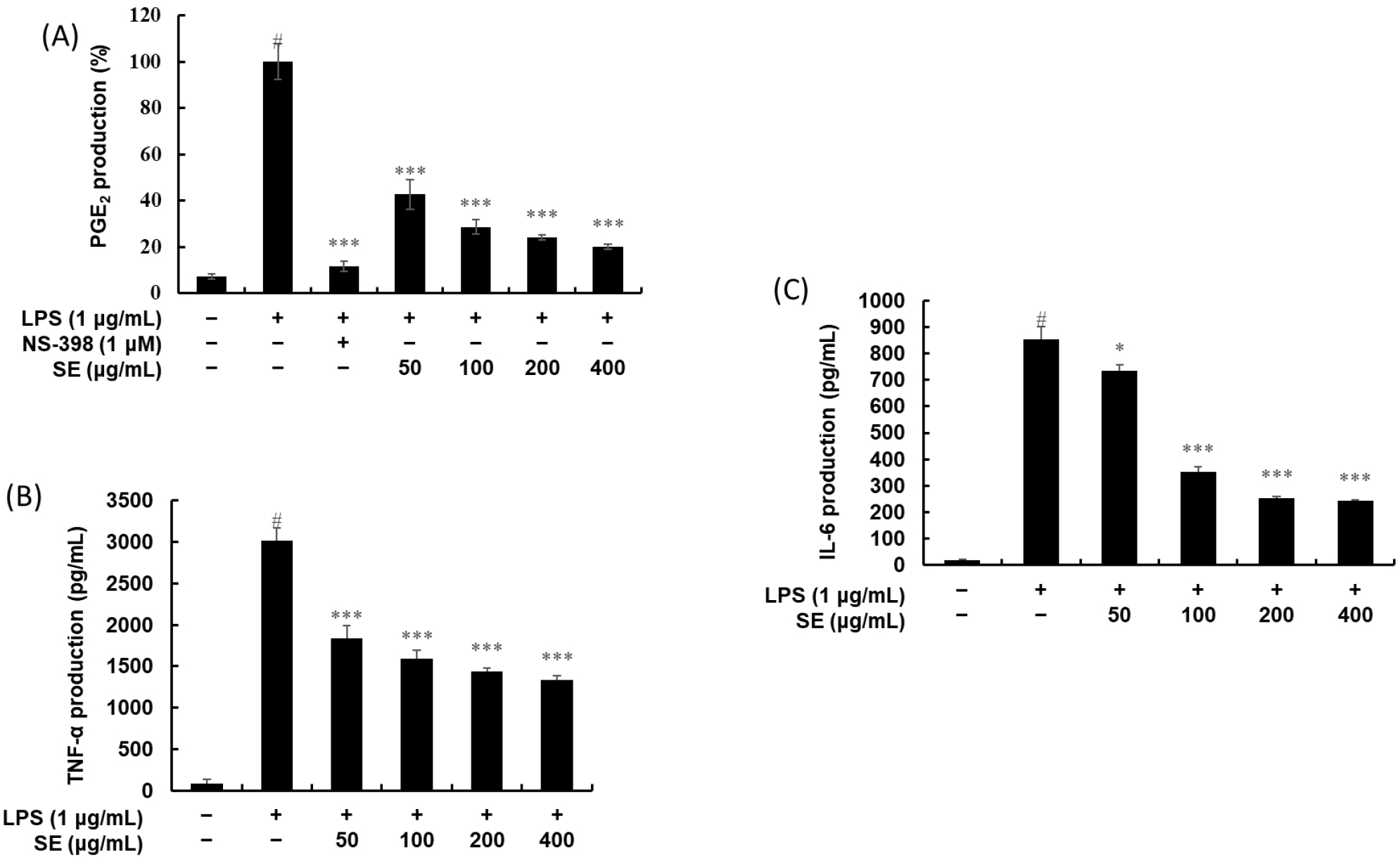
Cytokines, which are critical indicators of inflammation, are produced during the inflammatory process, signaling its progression. An ELISA kit was used to investigate the effect of SE on inflammatory cytokine production. Figs. 6B and 6C illustrate the anti-inflammatory effects of SE on two key cytokines, TNF-α and IL-6, respectively. In both graphs, unstimulated RAW 264.7 cells showed negligible production of TNF-α and IL-6. However, LPS treatment alone led to a marked increase in both cytokines, reaching over 3,000 pg/mL for TNF-α and over 800 pg/mL for IL-6. Subsequent treatment with increasing concentrations of SE (50, 100, 200, and 400 μg/mL) caused a significant and dose-dependent inhibitory effect on both TNF-α and IL-6 production. The inhibitory effect was particularly potent at the higher concentrations, demonstrating that SE effectively suppresses the inflammatory response by down-regulating these crucial cytokine mediators.
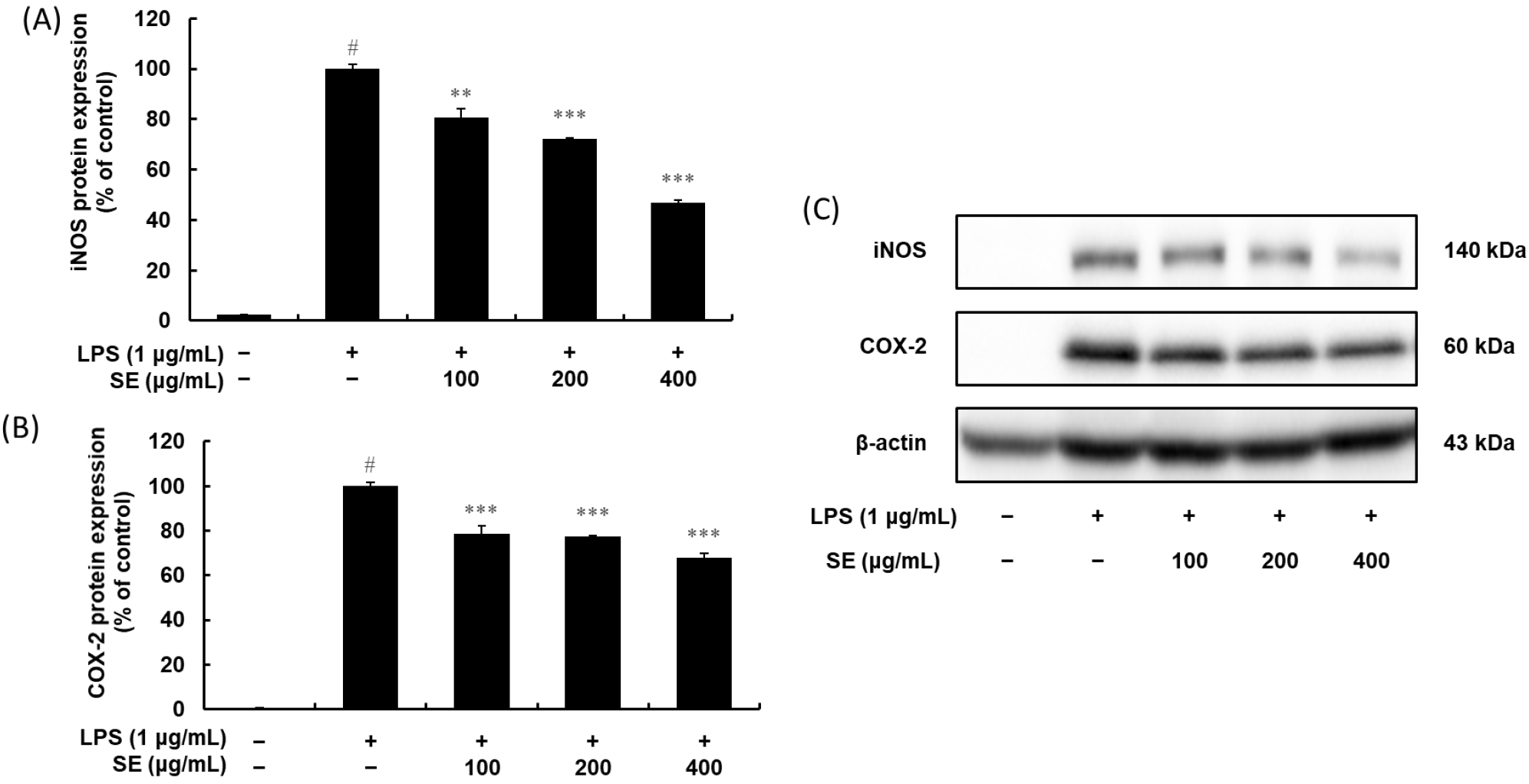
Given that NF-κB is a key regulator inducing both iNOS and COX-2, we examined how SE influenced the expression of these crucial inflammatory enzymes (Fig. 7). LPS treatment significantly increased iNOS and COX-2 protein expression, while simultaneously causing a decrease in cytoplasmic p65 protein as presented in section 3.4.1. Interestingly, SE treatment led to a dose-dependent increase in cytoplasmic p65, which prominently suppressed the LPS-induced iNOS and COX-2 protein expression. Thus, the observed reduction in iNOS and COX-2 protein expression is the downstream result of SE’s inhibitory control over the NF-κB pathway. Overall, this coordinated inhibitory action on the NF-κB pathway is the likely mechanism behind SE’s anti-inflammatory properties.
Besides the central transcription factor NF-κB, numerous inflammation-related genes are also regulated by MAPK-activated transcription factors (Fig. 8). To further understand SE’s anti-inflammatory mechanism, we examined its effect on the phosphorylation of ERK, JNK, and p38 MAPK in LPS-induced RAW 264.7 cells. We treated the cells with SE (100, 200, and 400 μg/mL) alongside LPS (1 μg/mL). While LPS stimulation of RAW 264.7 cells for 20 min robustly induced the phosphorylation of ERK, JNK, and p38 MAPK, SE significantly suppressed the phosphorylation of JNK and p38 MAPK in a dose-dependent manner. SE also showed a slight reduction in ERK MAPK phosphorylation in these LPS-activated cells. These findings indicate that citrus fruit extracts can effectively block MAPK signal transduction pathways in stimulated RAW 264.7 macrophages. Using Western blotting, we analyzed whole cell lysates to determine the maximum level of MAPK phosphorylation after LPS treatment. This was accomplished by using specific antibodies that bind to the phosphorylated forms of p38, ERK, and JNK.
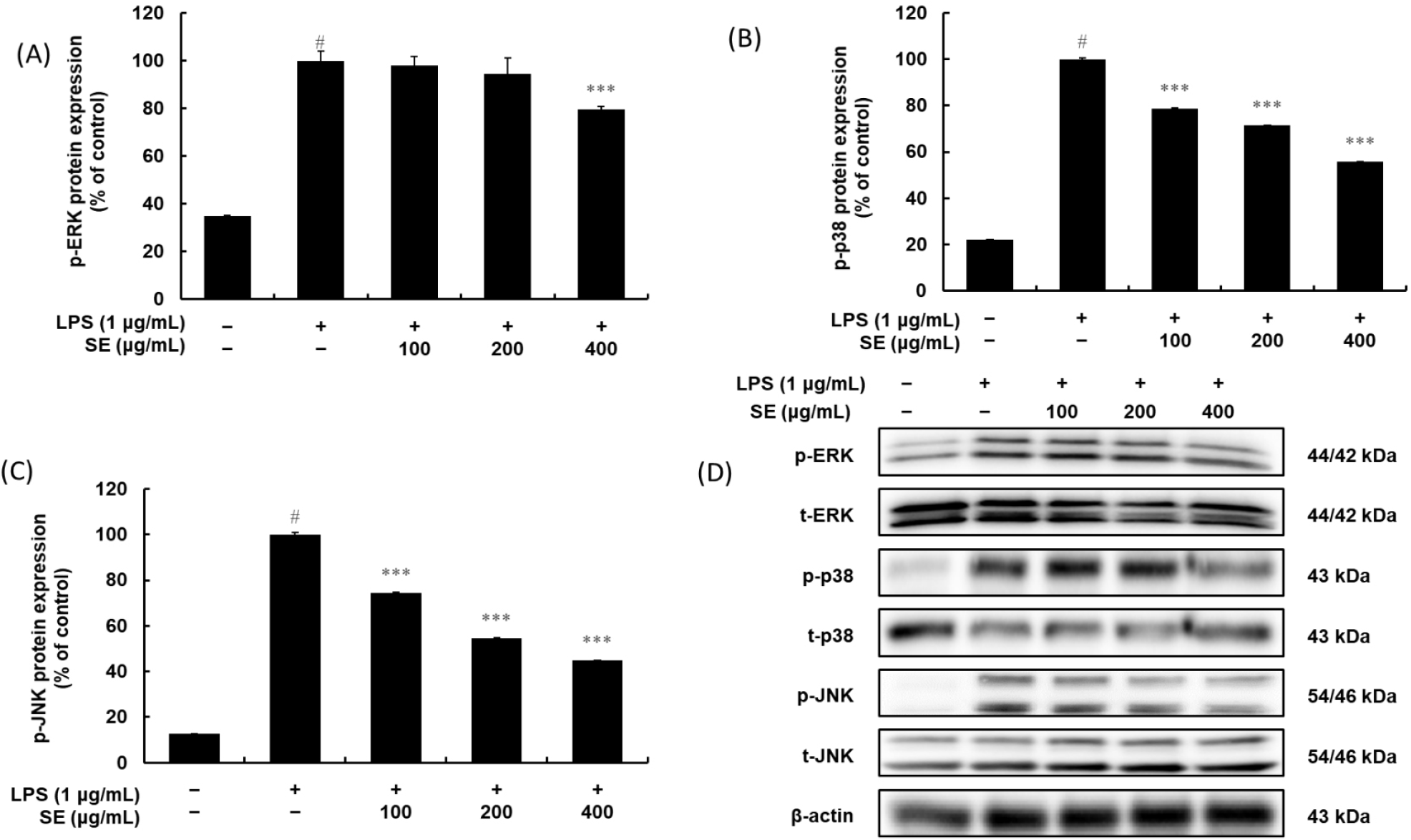
Anti-inflammatory agents also act on the MAPK signaling pathways, including ERK, p38, and JNK. These pathways are activated by LPS and can also regulate the activity of NF-κB and other transcription factors (Seger and Krebs, 1995). Inhibiting the phosphorylation of these kinases is another mechanism by which SE could dampen the inflammatory signal. In our study, SE significantly suppressed the phosphorylation of JNK and p38 MAPK in a dose-dependent manner while it slightly attenuated the ERK MAPK phosphorylation in these LPS-activated cells, indicating the inhibitory impact of the SE on LPS-induced cytokine production could also be explained by its ability to modulate MAPK signaling.
4. Conclusions
This study provides the first comprehensive time-course analysis detailing the concurrent evolution of quality parameters, organic acids, and five major flavonoids across the pulp and peel of a relatively new Sarahyang citrus variety. Our analysis reveals significant chemical shifts during maturation: sweetness, driven primarily by a consistent increase in sucrose (with correlated increases in fructose and glucose), was balanced by a significant decline in acidity, attributed to the sharp decrease in the dominant organic acid, citric acid. Crucially, the peel consistently emerged as the dominant repository for flavonoids, containing significantly higher concentrations than the pulp and serving as the exclusive source for the potent anti-inflammatory polymethoxyflavones, nobiletin and tangeretin.
This distinctive profile establishes the peel’s high industrial value, a finding critical for the valorization of this common food industry byproduct, as the fruit’s potent anti-inflammatory properties are intrinsically concentrated within these phytochemicals. Mechanistically, the SE demonstrated potent, dose-dependent anti-inflammatory action in LPS-stimulated RAW 264.7 cells. This multi-pronged inhibitory effect is achieved by blocking key inflammatory pathways: SE prevents NF-κB activation (by inhibiting IκB-α degradation) and suppresses the phosphorylation of both the JNK and p38 MAPK pathways. While the peel is the primary reservoir, the pulp and juice still contribute meaningfully to health-promoting activity through their bioactive content and potent antioxidative capacity.
Acknowledging the robust mechanistic depth achieved in vitro, a key limitation is that these findings are based on a murine macrophage cell line (RAW 264.7) and currently lack in vivo efficacy data. Therefore, future research should prioritize animal models to confirm the observed anti-inflammatory mechanisms, assess in vivo bioavailability, and evaluate safety. Subsequently, clinical trials focusing on inflammatory and metabolic markers are warranted to substantiate the health claims and guide the development of Sarahyang-based functional foods or nutraceuticals, particularly those utilizing the high-value peel byproduct.