
1. Introduction
Fermented foods are an important part of the Korean diet, and fermentation is involved in the production of many traditional foods (Shin, 2010). The quality of fermented foods is largely determined by microbial activity, which shapes both product safety and sensory attributes (Kim, 2011). Acetic acid bacteria (AAB) are Gram-negative, obligately aerobic bacteria that oxidize ethanol to acetic acid and play a central role in vinegar production. Because acetic acid productivity varies among strains and is influenced by their physiological characteristics, the selection of high-performing strains is critical for stable and efficient vinegar fermentation (Kim et al., 2023; Park et al., 2015). AAB are classified into several genera based on their oxidative metabolism. Acetobacter and Komagataeibacter are capable of oxidizing ethanol to acetic acid and further oxidizing acetic acid, whereas Gluconobacter preferentially oxidizes sugars and incompletely oxidizes alcohols, with limited ability to oxidize acetic acid. Representative species include Acetobacter aceti, A. pasteurianus, and A. liquefaciens, as well as Komagataeibacter xylinum and K. hansenii (Park et al., 2013).
Natural fermentation carries risks of contamination by exogenous microorganisms, leading to difficulties in process control and inconsistencies in taste, aroma, and color (Mun et al., 2018; Yang et al., 2017). The microbial diversity inherent in naturally fermented products often results in variable quality that is difficult to reproduce outside their place of origin (Chessa et al., 2023; N’Guessan et al., 2015). To overcome these limitations, the use of starter cultures has been proposed as an effective strategy to introduce appropriate microorganisms and alleviate spoilage problems during fermentation (N’Guessan et al., 2015). However, the Korean fermented food industry still largely relies on imported starter cultures (Kim, 2024). Therefore, the development of starter culture technology using indigenous microorganisms is essential and presents significant opportunities for improving the self-sufficiency of the Korean fermented food industry (Kim, 2023). While extensive studies have been conducted on dried starter cultures based on lactic acid bacteria and yeasts, research on AAB remains relatively limited due to their strict aerobic metabolism and sensitivity to environmental stresses (N’Guessan et al., 2015; Tsaousi et al., 2008; Utami et al., 2020). Thus, the development of dried starter cultures for AAB could further diversify vinegar production and related fermentation processes.
Previous studies have investigated the physicochemical characteristics of Acetobacter and Komagataeibacter species under various culture conditions (Baek et al., 2014; Mas et al., 2014; Mizzi et al., 2022). However, most of these studies have focused on characterizing individual strains under specific conditions, with limited research on starter culture development and industrial applications. Therefore, this study aimed to compare the acetic acid production and volatile compound profiles of ten AAB strains originating from Korean traditional fermented vinegars, as well as to investigate optimal culture conditions for cell growth and acetic acid production. Based on these results, suitable strains were selected, followed by evaluation of the quality characteristics of their dried starter cultures.
2. Materials and methods
Ten AAB strains (Table 1) were obtained from the Korean Agricultural Culture Collection (KACC). These strains were previously isolated from Korean traditional vinegars and identified by the provider using 16S rRNA gene sequencing according to standard procedures. In this study, the strains were stored at -70°C and cultivated on SM agar (yeast extract 0.5%, glucose 3.0%, CaCO3 1.0%, agar 2.0%, ethanol 5.0%, w/v) at 30°C. For activation, single colonies were inoculated into LM broth (yeast extract 0.5%, glucose 0.5%, glycerin 1.0%, MgSO4 ․ 7H2O 0.02%, ethanol 5.0%, acetic acid 1.0%, w/v) and incubated at 30°C with shaking at 150 rpm for 72 h.
To compare the growth of the ten AAB strains, 1 mL of culture broth was diluted to an appropriate concentration, and the optical density was measured at 600 nm using a spectrophotometer (Cary 60 UV-Vis, Agilent Co., Santa Clara, CA, USA). pH was measured using a pH meter (Orion 3 star, Thermo Fisher Scientific, Waltham, MA, USA). Total acidity was determined by diluting 1 mL of the sample with 9 mL of distilled water, using phenolphthalein as an indicator, and titrating with 0.1 N NaOH to pH 8.2, followed by conversion of the titration value (mL) into acetic acid equivalents (%). To verify that acetic acid was the major contributor to total acidity, acetic acid concentration was quantified by HPLC. Samples were filtered through a 0.2 μm membrane filter (Millipore, Co., Cork, Ireland) and analyzed using an HPLC system (LC-20A, Shimadzu Co., Kyoto, Japan) equipped with a TSKgel ODS-100V column (5 μm, 4.6×250 mm, Tosoh Co., Tokyo, Japan) and a UV detector (440 nm). The mobile phase consisted of 8 mM perchloric acid (mobile phase A) and a mixture of 0.2 mM bromothymol blue, 15 mM Na2HPO4, and 7 mM NaOH (mobile phase B). The analysis was conducted at 40°C with an isocratic mobile phase at a flow rate of 1 mL/min, and an injection volume of 10 µL.
For electronic nose analysis, 3 mL of each sample was placed in a 20 mL vial and analyzed five times using an electronic nose (Heracles NEO, Alpha MOS, Toulouse, France). Two columns (MTX-5 and MTX-1701) were operated in parallel and coupled with flame ionization detectors (FID). The analysis conditions were set as follows: injection volume 3 mL, column temperature 50°C, trapping temperature 40°C, injection temperature 200°C, FID temperature 260°C, and isocratic flow rate 1.0 mL/min. Aroma pattern analysis was performed using principal component analysis (PCA) with Alpha MOS software.
To determine optimal growth conditions for biomass production, the effects of culture medium composition, temperature, aeration, carbon sources, and nitrogen sources were evaluated. Three culture media were tested: AE medium (yeast extract 0.3%, peptone 0.4%, glucose 0.5%, acetic acid 3.0%, and ethanol 3.0%), GYE medium (glucose 5.0%, yeast extract 1.0%, and ethanol 1.0%), and ethanol-based LM medium (yeast extract 0.5%, glucose 0.5%, MgSO4 ․ 7H2O 0.02%, ethanol 6.0%, acetic acid 1.0%). Cultivation temperatures (20, 25, 30, and 35°C) and aeration conditions (static, 100 rpm, and 250 rpm) were examined. To evaluate nutrient effects, sucrose, fructose, lactose, galactose, and maltose were tested as carbon sources, while tryptone, (NH4)2SO4, NH4Cl, NaNO3, and KNO3 were evaluated as nitrogen sources. Cell growth and acid production were assessed by measuring optical density at 600 nm and total acidity (%), respectively.
Fed-batch cultivation was conducted in a 5 L jar fermenter (KoBioTech Co., Ltd, Incheon, Korea) at 30°C with an agitation speed of 600 rpm. The pH was maintained at 6.0, and dissolved oxygen (DO) was controlled at saturation levels. The modified medium contained 0.5% yeast extract, 0.5% glucose, 1.0% glycerin, 0.02% MgSO4 ․ 7H2O, and 6% ethanol. Ethanol (3%) was intermittently supplemented to maintain its concentration, and antifoam was added to suppress foam formation. During fed-batch cultivation, metabolite profiles, including ethanol consumption and organic acid production, were monitored by HPLC. Samples were filtered through a 0.2 μm membrane filter and analyzed using an HPLC system (Hitachi 5000 Chromaster, Hitachi Ltd, Tokyo, Japan) equipped with an Aminex HPX-87H column (300×7.8 mm, Bio-Rad Laboratories, CA, USA) and a refractive index detector (RID). The mobile phase was 0.005 N sulfuric acid, with a column temperature of 50°C, a flow rate of 0.6 mL/min, and an injection volume of 10 μL.
For starter culture production, cells were harvested by centrifugation, suspended in 20% skim milk as a cryoprotectant, and lyophilized. The dried cells were blended with maltodextrin, lactomil (89% lactose, 11% maltodextrin), β-cyclodextrin, and skim milk as drying carriers at a ratio of 0.1% (w/w, dried cells to carrier), targeting a viable cell count of 7.00 log CFU/g.
Moisture content was determined using the atmospheric drying method. One gram of sample was dried at 105°C using a moisture analyzer (MX-50, A&D Ltd, Tokyo, Japan) until a constant weight was reached. Water activity (Aw) was measured using an Aw analyzer (LabPartner-Aw, Novasina AG, Switzerland) at room temperature. Viable cell counts were determined by serial dilution in sterile distilled water and plating on SM agar. Plates were incubated at 30°C for 96 h, and colonies were counted as log CFU/g. AAB colonies were identified by the formation of clear zones due to CaCO3 dissolution.
Alcoholic fermentation broth was prepared using nuruk and Saccharomyces cerevisiae YM45 through sequential fermentation at 25°C to obtain an ethanol-containing substrate. The broth was adjusted to 6% (v/v) ethanol and inoculated with 0.25% (w/v) dried starter cultures stored at 4°C for five weeks. Vinegar fermentation was conducted under static conditions at 30°C for 168 h. Acetic acid concentration was determined by HPLC as described above.
All results are presented as mean±SD. Statistical analyses were performed using the statistical analysis system SPSS version 27 (IBM Corp., Armonk, NY, USA). For significant differences between each group, t-test or one-way analysis of variance was performed at the p<0.05 level, and significant differences were verified using Duncan’s multiple range test.
3. Results and discussion
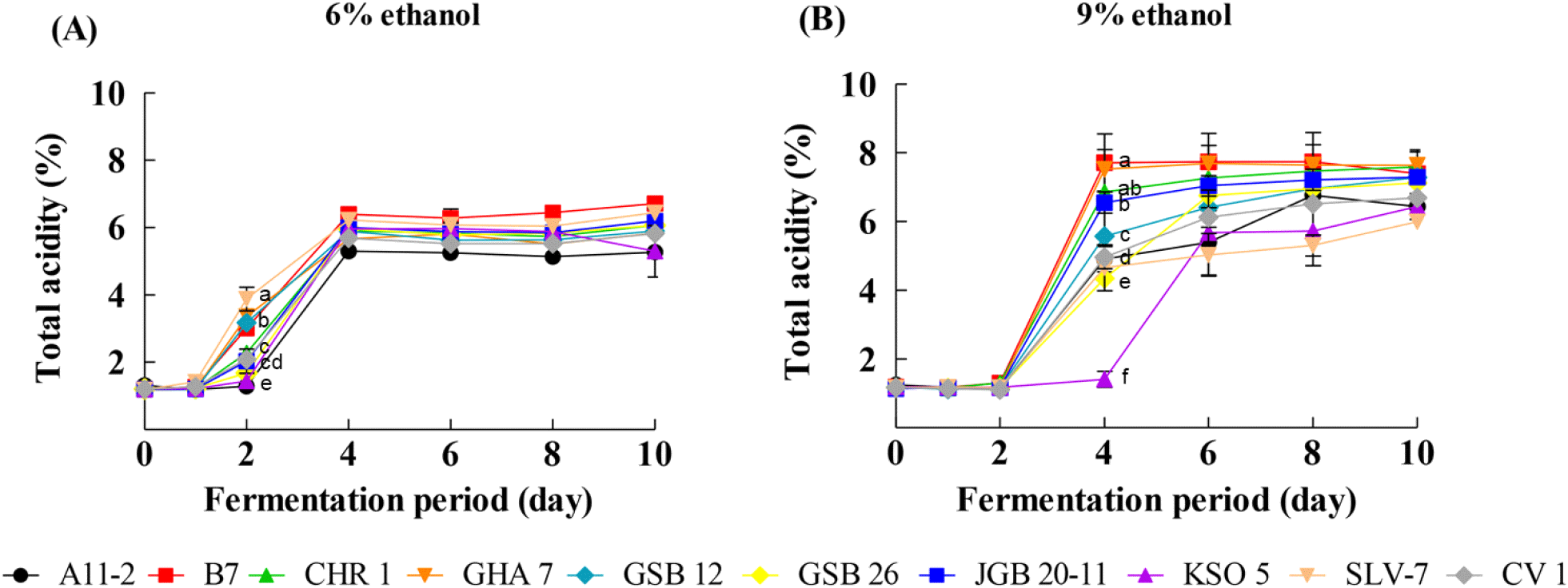
Variations in total acidity and acetic acid production among the ten AAB strains are shown in Fig. 1. Under 6% initial ethanol conditions, several strains, including A. oryzifermentans SLV-7 and A. pasteurianus isolates, produced high total acidity (5.91-6.39%), indicating efficient oxidation of ethanol to acid (Fig. 1A). At 9% initial ethanol conditions, A. pasteurianus B7, JGB 20-11, GHA 7, GSB 12, and CHR 1 exhibited increased acid production (6.75-7.74%) (Fig. 1B). HPLC analysis confirmed high acetic acid production in A. pasteurianus B7 (83.2±6.3 g/L) and G. saccharivorans CV 1 (83.1±12.5 g/L), indicating that most of the total acidity was attributable to acetic acid (Table 2). Previous studies have reported that increasing the initial alcohol concentration generally reduces conversion efficiency (Cho et al., 2015). Since AAB produce acetic acid through ethanol oxidation, the initial ethanol concentration is a critical factor affecting both microbial growth and acid production (Sim et al., 2001). Excessively high ethanol concentrations may inhibit cell growth and decrease acid productivity (Lee et al., 2023). In contrast, most strains in this study maintained high acetic acid production even at 9% ethanol, indicating strong alcohol tolerance and fermentation efficiency. These results suggest that the selected strains are suitable candidates for high-acidity vinegar production.
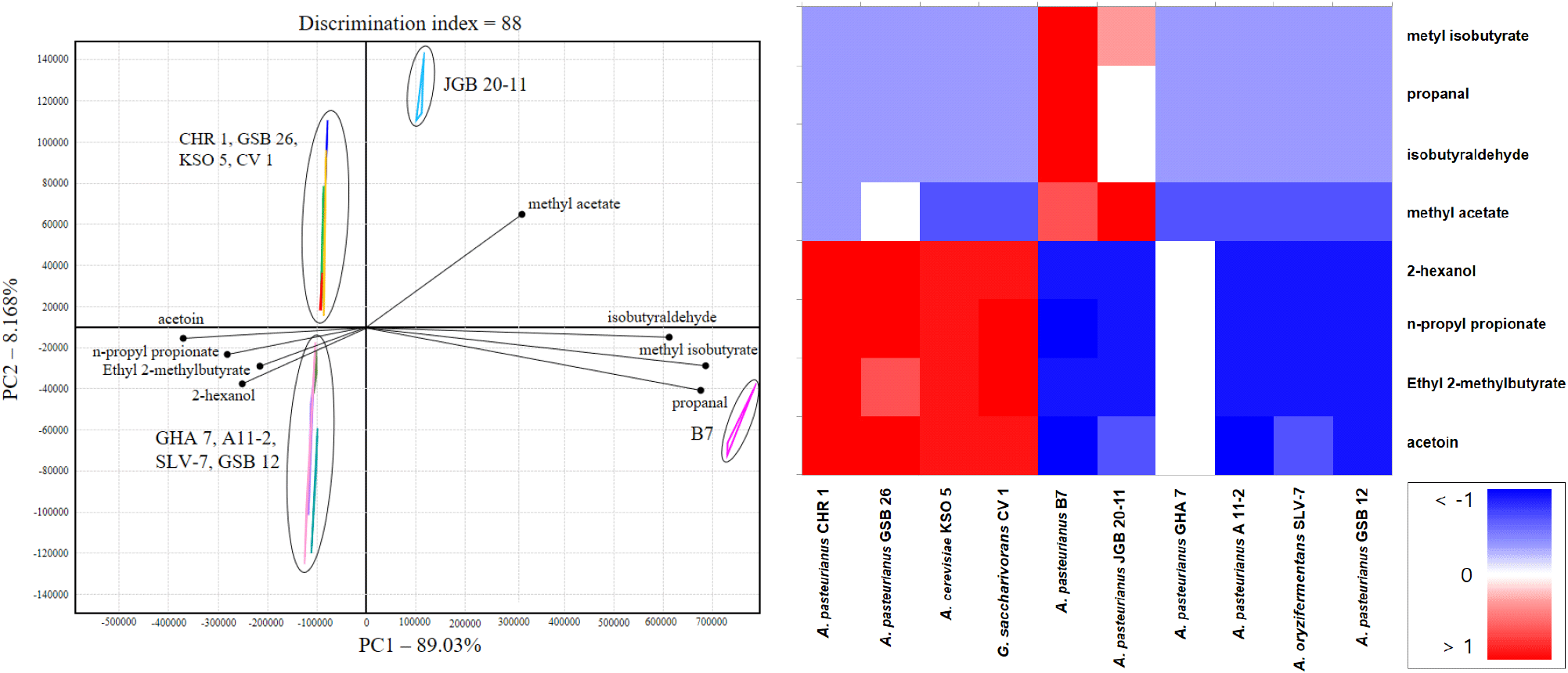
Electronic nose technology enables characterization of aroma properties by simulating human sensory perception and allows comparative analysis of volatile profiles among samples (Jiang et al., 2025). The electronic nose results for the ten AAB strains are presented in Fig. 2A. PCA explained 97.19% of the total variance (PC1: 89.03%, PC2: 8.16%). Along the PC1 axis, A. pasteurianus B7 and JGB 20-11 were positioned in the positive direction, indicating distinct aroma patterns compared with the other strains. Volatile compounds with chromatogram values exceeding 1,000 and discrimination power greater than 0.8 are shown in Fig. 2B. Eight compounds were identified: methyl acetate, methyl isobutyrate, propanal, isobutyraldehyde, ethyl 2-methyl butyrate, acetoin, 2-hexanol, and n-propyl propionate. Methyl acetate is associated with fruity and floral notes (Kolby et al., 2014; Maia et al., 2021), whereas ethyl 2-methyl butyrate contributes floral note and methyl isobutyrate contributes floral and fresh notes (Jiang et al., 2025). The chromatogram value of methyl acetate was 5,435.6±722.1 for A. pasteurianus B7, 13,573.1±76.0 for A. pasteurianus JGB 20-11, and 1,215.5±107.5 for A. pasteurianus GSB 26. For methyl isobutyrate, the values were 74,477.9± 4,968.1 for A. pasteurianus B7, 14,111.1±1,360.8 for A. pasteurianus JGB 20-11, and 2,688.8±117.4 for A. pasteurianus GHA 7. For isobutyraldehyde, the value was 1,328.1±381.2 for A. pasteurianus B7. These results suggest that certain strains produce desirable fruity and floral aromas, making them suitable for enhancing flavor in vinegar. A. pasteurianus B7 and A. pasteurianus JGB 20-11 exhibited comparable acid production levels and distinct volatile patterns compared with the other strains and were therefore selected as candidate strains. In addition to their distinct volatile profiles, both strains exhibited high acetic acid production under elevated ethanol conditions, suggesting their suitability for efficient industrial vinegar fermentation. Moreover, A. pasteurianus is a species with a long history of safe use in vinegar production, and the origin of the selected strains from traditional vinegars further supports their safety and applicability as starter cultures.
To establish culture conditions suitable for biomass production of A. pasteurianus B7 and A. pasteurianus JGB 20-11 as dried starter candidates, the effects of carbon sources, nitrogen sources, aeration conditions, and temperature on cell growth were evaluated (Fig. S1-S4). The tested conditions were consistent with those reported in previous studies (Kim et al., 2023; Park et al., 2002; Raspor and Goranovič, 2008; Sainz et al., 2017). Cell growth was monitored as OD600 during cultivation, and these screening experiments were conducted to identify conditions that support rapid biomass accumulation prior to scale-up fermentation. Across the carbon sources tested, sucrose supported the highest growth for both strains; however, most alternative carbon sources produced similar growth trends, indicating that these strains can grow well without a pronounced loss of biomass production. In contrast, yeast extract consistently promoted the most robust growth compared with the other nitrogen sources evaluated, suggesting that complex nutrients and growth factors are advantageous for rapid biomass accumulation. Since AAB growth is strongly oxygen-dependent, aeration was also critical. Agitation at 250 rpm (aerobic, highly agitated conditions) yielded the greatest cell growth relative to lower agitation or static conditions. Temperature screening further showed that 30-35°C was optimal for maximizing growth. Both strains still exhibited measurable growth even at 20°C, indicating a temperature tolerance that may be advantageous for vinegar production under practical conditions where strict temperature control is difficult.
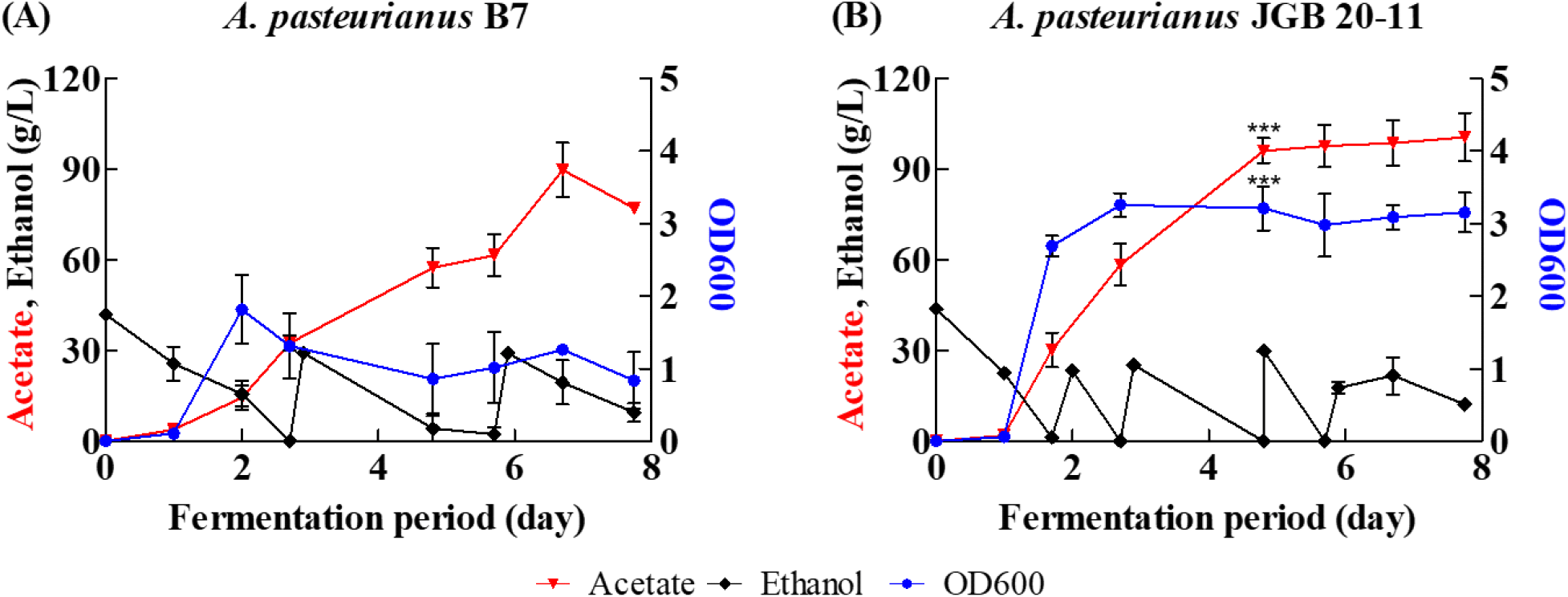
Fed-batch cultivation of A. pasteurianus B7 and A. pasteurianus JGB 20-11 was carried out to achieve high biomass production and acetic acid formation by maintaining the ethanol concentration at approximately 30 g/L. Fermentation profiles of the fed-batch cultivation are shown in Fig. 3. A. pasteurianus B7 produced 89.74±12.72 g/L acetic acid after 160 h of fermentation, and the maximum OD600 of 1.82± 0.67 was reached after 24 h of fermentation (Fig. 3A). A. pasteurianus JGB 20-11 produced 100.59±11.35 g/L acetic acid after 186 h of fermentation, and the maximum OD600 of 3.26±0.23 was reached after 65 h of fermentation (Fig. 3B). In both cases, lactate, glycerol, and other by-products were detected at negligible levels, indicating that carbon flux was efficiently directed toward acetic acid production. For both strains, biomass production and acetic acid formation were enhanced compared with flask cultivation, demonstrating that the fed-batch strategy is suitable for producing highly concentrated AAB biomass as a precursor for dried starter cultures.
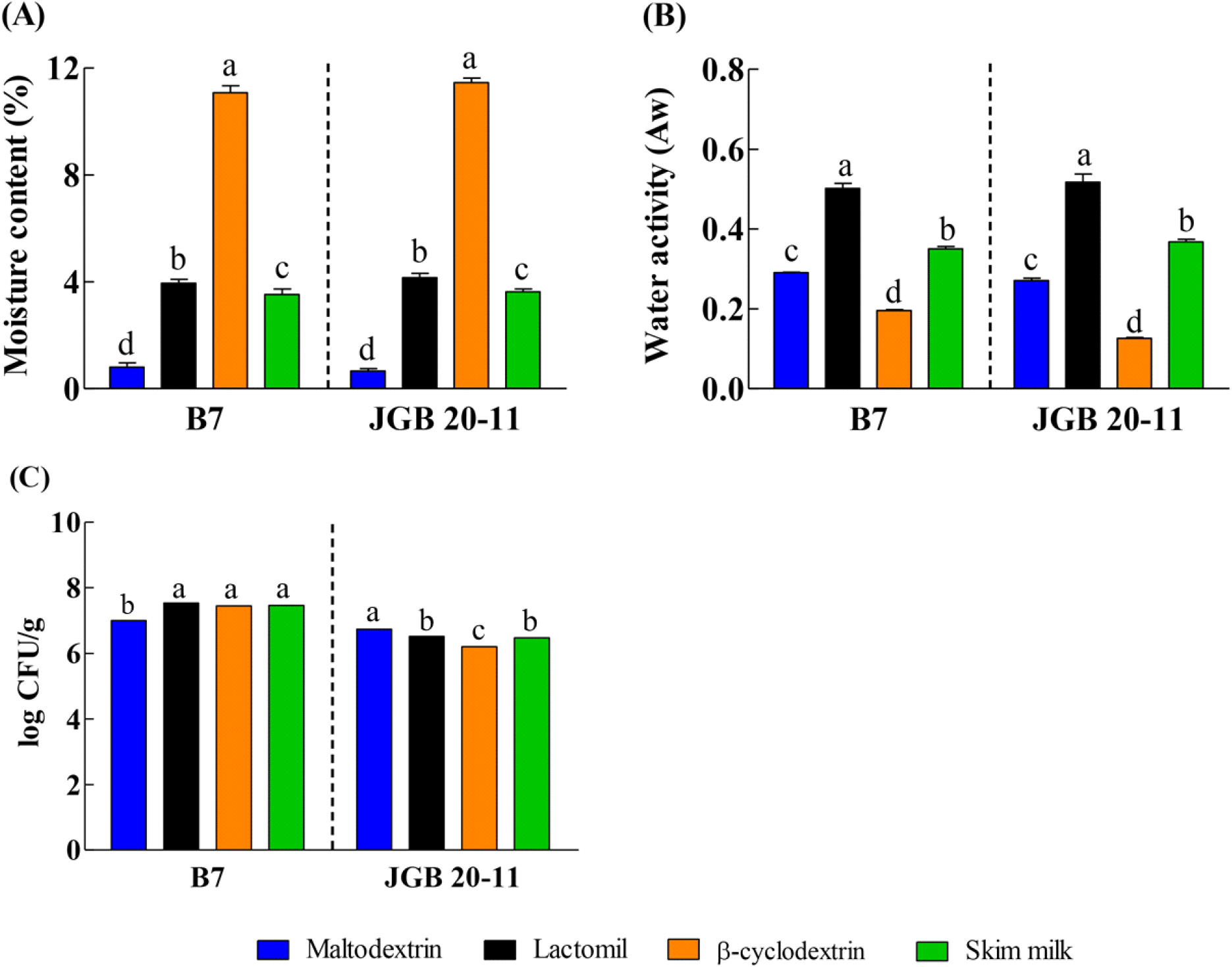
A. pasteurianus B7 and A. pasteurianus JGB 20-11 were lyophilized and blended with various drying carriers. Moisture content and Aw are important properties influencing the storage stability of dried materials (Lima et al., 2025). Low moisture content (<6%) and Aw values (<0.6) are generally regarded as practical limits for suppressing microbial growth and limiting deterioration reactions, thereby contributing to shelf-life stability of particulate products (Lima et al., 2025; Ravichandran, 2023). The moisture content of dried starter cultures blended with maltodextrin, lactomil, and skim milk ranged from 0.80% to 3.95% (Fig. 4A), and the Aw values of all dried starter cultures were below 0.6 (Fig. 4B). Although β-cyclodextrin exhibited a relatively low Aw, it showed a higher moisture content compared with the other drying carriers tested. Since all drying carriers exhibited Aw values below 0.6, these results suggest potential microbiological stability of the dried starters under the tested conditions. Furthermore, considering potential moisture ingress during storage and distribution, drying carriers associated with lower moisture content may be more favorable. Overall, these results indicate that maltodextrin, lactomil, and skim milk can function as effective drying carriers for producing dried starter cultures with microbiological and chemical stability (Shishir and Chen, 2017). In addition, microbiological quality is a critical factor for starter cultures, as excessive contamination may lead to spoilage of fermented products. In this study, AAB starter cultures prepared with different drying carriers maintained viable cell counts of approximately 7 log CFU/g immediately after drying (Fig. 4C). This level of viability is comparable to the cell density ranges reported in previous studies for AAB populations observed during vinegar fermentation or starter-related preparations (Sengun et al., 2022). These findings indicate that the dried cultures retained a level of viable cells comparable to those reported in previous studies. However, maintaining cell viability alone does not guarantee fermentation performance; therefore, their functional efficacy was further evaluated in vinegar fermentation. When the dried starter of A. pasteurianus B7 blended with lactomil was applied to vinegar fermentation, complete ethanol consumption was achieved within 7 days, producing 48.13±1.25 g/L acetic acid, indicating effective fermentation performance (Fig. 5). Although the other formulations also consumed ethanol, acetic acid production was comparatively lower.
4. Conclusions
Acetic acid bacteria strains isolated from Korean traditional vinegars were evaluated as starter cultures for high-acidity vinegar production. Among the ten isolates, A. pasteurianus B7 and A. pasteurianus JGB 20-11 exhibited high acetic acid production under both 6% and 9% initial ethanol conditions and were characterized by fruity and floral volatile compounds, indicating efficient ethanol oxidation, alcohol tolerance, and favorable flavor profiles. Fed-batch cultivation enabled high cell density and acetic acid production with negligible formation of by-products such as lactate and glycerol. The biomass of the selected strains was converted into dried starter cultures using maltodextrin, lactomil, and skim milk as drying carriers, resulting in low moisture content and low water activity (Aw), which indicate favorable physicochemical stability. Overall, A. pasteurianus B7 and A. pasteurianus JGB 20-11 are promising indigenous AAB starter strains, and the scale-up and drying processes established in this study provide a practical basis for industrial starter culture production and for improving the quality and consistency of vinegar.