
1. Introduction
Cassava (Manihot esculenta Crantz) is one of the most important staple crops that contribute to food and nutritional security in sub-Saharan Africa (Adebayo, 2023). It is an important source of calorie, protein, micronutrients and bioactive compounds (Bayata, 2019; Zekarias et al., 2019). Nigeria, being the largest producer of cassava accounts for 63 million tonnes in 2023, representing 20% of global supply (Otekunrin, 2024). Globally, cassava utilization is largely for food and it is increasingly used to substitute wheat and other cereals in composite flours and feeds (Vilanculos et al., 2024). However, greater proportion of cultivated cassava is subjected to either solid-state or submerged fermentation before consumption in sub-Saharan Africa. They are processed into various traditional fermented foods, such as agbelima, atieke, fufu, gari, lafun, kivunde, pupuru and togwa (Adebayo, 2023; Banwo et al., 2020; Bouatenin et al., 2019).
Spontaneous fermentation of cassava into various food products is mediated by the most adapted microorganisms from the microflora of cassava and contaminating microorganisms from environmental sources. The predominant microbial groups include lactic acid bacteria (LAB) and yeasts. They contribute to microbiological safety, detoxification of cyanogenic glucosides, nutritional enrichment, flavour development and retting in/on the derived cassava products (Abimbola et al., 2024; Aghemwenhio et al., 2017a; Edward et al., 2012; Ogunremi and Sanni, 2011; Oyewole, 2001).
The most important limitation to the utilization of cassava for food is the presence of toxic cyanogenic glucosides (linamarin and lotaustralin) (Bouatenin et al., 2019; Edward et al., 2012). Upon consumption of poorly processed cassava, these compounds are implicated in acute cyanide poisoning, with signs and symptoms including nausea, vomiting, diarrhea, dizziness and weakness (Kashala-Abotnes et al., 2019; Otun et al., 2023; Quinn et al., 2022). Long-term exposure results to chronic cyanide poisoning, leading to goiter, cretinism, retarded growth, neurological defects such as konzo and tropical ataxic and sometimes death (Kashala-Abotnes et al., 2019; Quinn et al., 2022).
Previous reports have demonstrated the potentials of yeasts to contribute to the detoxification of cassava by producing β-glucosidase. The enzyme breaks down cyanogenic glucosides during fermentation. Strain-specific β-glucosidase production was reported in some species of Candida from fermenting cassava mash for fufu and gari production (Edward et al., 2012; Oyewole, 2001). Besides the importance of β-glucosidase-producing yeasts in cassava fermentation, they may offer the benefits of in vivo degradation of cyanogenic glucosides, if they possess probiotic traits.
Probiotics are live microorganisms that, when administered in adequate amounts, confer a health benefit on the host (Hill et al., 2014). Prospective probiotic microorganisms must demonstrate potentials to tolerate harsh conditions during gastrointestinal transit and persist in the intestine (Wang et al., 2024). Yeasts with probiotic potentials have been characterized from fermented foods produced from different substrates, including milk, meat, cereals, vegetables, juices and saps (Shruthi et al., 2022; Staniszewski and Kordowska-Wiater, 2021). To date, there are limited studies on the probiotic potentials of yeasts isolated from cassava-based fermented food products.
Fufu is a major part of the diet of consumers in Nigeria and some sub-Saharan African countries. It is a thick fine paste that is obtained by submerged fermentation of cassava. The cassava is peeled, washed, cut and steeped in water for 4-5 days. The retted cassava is then sieved, and allowed to sediment before decantation (Abimbola et al., 2024; Banwo et al., 2020). Gari is the most important cassava fermented product that is consumed across West Africa. It is a white or creamy, sour, and gritty product that is obtained by fermenting, dewatering and roasting fresh cassava mash (Banwo et al., 2020). Both fufu and gari are unexplored reservoir of yeasts that combine cyanogenic glucosides-degrading with probiotic potentials. Therefore, in this study yeasts were isolated from fufu and gari and characterize for technological and probiotic potentials, with the aim of developing detoxified and potentially healthful cassava-based fermented product.
2. Materials and methods
A total of 17 samples of cassava mash for fufu (9 samples) and gari (8 samples) production respectively were aseptically collected in sterile containers from clusters of artisanal producers in Ibadan, Nigeria. The traditional processing of fufu and gari are described by Banwo et al. (2020). They were immediately transported to Food Microbiology Laboratory, University of Ibadan, for analyses.
pH was measured using a calibrated pH meter (Corning Pinnacle M530 pH meter, Corning Inc., Corning, NY, USA) after homogenizing 40 g of each fermented food sample in 40 mL of sterile distilled water with a sterile blender (Ninja C5, Massachusetts, USA) (Edward et al., 2012).
Ten grams of each fermenting cassava mash sample was weighed and serially diluted into 90 mL of sterile saline water (0.85% NaCl). Tenfold serial dilutions of each sample were obtained and aliquots (1 mL) of appropriate dilutions were separately pour-plated in Potatoes Dextrose Agar (PDA™ Media, Delhi, India), Malt Extract Agar (MEA™ Media, Delhi, India) and Yeast Extract Peptone Dextrose Agar (YPDA™ Media, Delhi, India), which were supplemented with 200 mg/L of streptomycin sulphate (Sigma-Aldrich, St. Louis, Missouri, USA) to inhibit bacterial growth. The plates were incubated at 30°C for 48 h and the colonies formed were counted (Kurtzman et al., 2011). Distinct colonies were randomly selected and purified by repeated streaking on MEA, without streptomycin sulphate. Pure cultures were preserved on MEA agar slants at 5°C and renewed at 2-weekly intervals (Kurtzman et al., 2011).
Standard methods were used to determine the phenotypic characteristics of the yeast isolates. The tests carried out include; sugar fermentation, hydrolysis of urea, gelatin liquefaction, acid production from glucose, and tolerance to 1% acetic acid (Kurtzman et al., 2011). Molecular characterization of a selected yeast isolate was carried out by extracting the genomic DNA and amplifying the ITS region of the gene using universal primers; 5’-TCCGTAGGTGAACCTGCGG-3’ and 5’-TCCTCCGCTTATTGATATGC-3’ for ITS-1 and ITS-4 respectively (White et al., 1990). The amplification was performed under conditions described by Bautista-Gallego et al. (2011). The amplicon was sent to Inqaba Biotech, Ibadan, Nigeria for sequencing and the obtained sequence was aligned to a nonredundant database (NCBI, 2025) to identify the yeast isolate.
Yeast isolates were incubated on MEA at 30°C for 48 h. The colonies were dispensed using sterile loop into sterile saline water (0.85% NaCl) to have a turbidity that compare with McFarland Standard No. 3. Amylase and protease production were determined as described by Oyewole (2001) and Ogunremi et al. (2015) respectively. β-glucosidase production was determined as the ability of yeast isolates to release p-nitrophenol from p-nitrophenyl-β-D-glucopyranoside (PNPG) (Targetmol, Boston, USA) a synthetic analogue of linamarin as described by Edward et al. (2012). Briefly, PNPG medium was prepared by adding 0.1 g of PNPG to 100 mL of 0.666 M NaH2PO4 (pH 6). The mixture was dissolved and filter-sterilized using 0.45 μm pore sized syringe filter (Sartorius, Gottingen, Germany). Exactly 0.75 mL of yeast cell suspension was added to 0.25 mL of the PNPG medium and incubated at 30°C for 12 h. Formation of distinct yellow color indicates the degradation of PNPG and release of p-nitrophenol due to β-glucosidase production. The intensity of the yellow coloration was graded as light yellow, moderate yellow and deep yellow.
Strong β-glucosidase-producing yeast isolates were selected and evaluated for their tolerance to some simulated stress factors during cassava fermentation by measuring their growth in 5 mL of sterile YPD broth supplemented with either 800 mg/L of filter-sterilized potassium cyanide (KCN) (Ogbonnaya, 2016), lactic acid (0.6% and 1.2%) or acetic acid (0.15% and 0.30%) (Perricone et al., 2014). The use 800 mg/L of filter-sterilized potassium cyanide (KCN) represents the extreme upper limits reported for some cassava varieties cultivated in cases of high temperature, drought and CO2 pressure (Cardoso et al., 2005; Nyaika et al., 2024). The same concentration of KCN was also used to screen for cyanide tolerant yeasts by Ogbonnaya (2016). The YPD broths were inoculated with 1% (v/v) of 24 h old broth culture of each yeast isolate (106 CFU/mL) and incubated at 30°C for 24 h. Unsupplemented YPD broth culture served as positive control. Yeast growth was determined by measuring the absorbance at 600 nm (Perricone et al., 2014). The tests were carried out in triplicates and growth index (GI) was calculated using the formulae;
where Abss was the absorbance of the respectively supplemented YPD broth culture, whereas Absc was the absorbance of the untreated YPD broth culture (positive control) (Perricone et al., 2014).
Fresh broth cultures of selected yeasts were washed in phosphate buffered saline (PBS) (0.1 M; pH 7.2). The yeast suspensions in PBS were standardized to 106 CFU/mL and used for the in vitro determination of probiotic potentials based on the following tests.
Survival of the yeast cells at low pH (2.5 and 3.5), following incubation at 37°C for 3 h and different concentrations of bile salts (0.3 and 0.5%) at 37°C for 24 h was determined as described by Ogunremi et al. (2015). Percentage survival was calculated;
where CFU/mL (Final) was the viable count of yeast after respective incubation time, and CFU/mL (Initial) was the viable count of yeast before incubation.
Conditions in the stomach and intestine were simulated and survival during gastrointestinal tract passage was evaluated as described by Pennacchia et al. (2008). Percentage survival at respective stage of the passage was calculated using the formula in subsection 2.6.1.
Exactly 3 mL of standardized yeast cell suspension was measured in sterile test tubes. The absorbance (600 nm) of the suspension was determined and equal volume of toluene and xylene were added separately to the cell suspension. The mixture was vortexed for 2 min, and left to stand for 45 min at 37°C to allow for phase separation. The aqueous phase was carefully obtained and the absorbance was determined at 600 nm (Van Holle et al., 2012). The adherence to respective hydrocarbon was calculated as hydrophobicity according to the equation below:
where A0 is the initial absorbance of the cell suspension, and A is the absorbance of the aqueous phase.
Exactly 10 mL of standardized yeast cell suspension was measured in sterile test tubes. The suspensions were vortexed for 10 sec and incubated at 37°C for 24 h. Two mL of the upper suspension was carefully taken at 2, 4 or 24 h and the absorbance was determined at 600 nm (Ogunremi et al., 2015). The auto-aggregation percentage was expressed as;
At represents the absorbance at time t = 2, 4 or 24 h; A0 represents the absorbance at 0 h.
Intracellular cell-free extracts obtained from selected test yeasts were prepared as described by Ogunremi et al. (2015). The antioxidant activity was analyzed, following 1,1-diphenyl-2-picryl-hydrazyl (DPPH) scavenging assay (Sharma and Bhat, 2009). Exactly 1 mL of 200 μM DPPH in methanol was mixed with 1 mL of each intracellular cell-free extract. In a control reaction tube, 1 mL of DPPH solution was mixed with 1 mL of phosphate buffer. The reaction tubes were maintained in the dark using aluminum foil and kept at 30°C for 30 min. The absorbance was recorded at wavelength of 517 nm. The percentage of antioxidant activity was derived using the following formula:
A0: Absorbance of control, A: Absorbance of intracellular cell-free extract.
Cassava mash was subjected to controlled fermentation using selected yeast strain with β-glucosidase-producing, stress tolerance and probiotic potentials. Yeast inoculum was prepared by culturing (30°C/24 h) selected strain in YPD broth and underwent two washes with equivalent volume of sterile saline solution (0.89% NaCl) (Ogunremi et al., 2022). The yeast cells were standardized to 107 CFU/mL. Exactly 100 g of peeled cassava was surface-sterilized (96% ethanol) and aseptically peeled, washed, chopped and steeped in 500 mL of sterile distilled water. The steeped cassava was inoculated with 1% (v/v) of standardized yeast cell suspension to start the fermentation process at 28-30°C for 72 h. An uninoculated stepped cassava model was used as negative control (Oyewole, 2001). Samples were aseptically taken at 24-hourly intervals for determination of yeast counts (Kurtzman et al., 2011), pH (Edward et al., 2012), and residual cyanide content (Halake et al., 2019). Briefly, alkaline picrate method was used to measure the residual cyanide levels of the fermenting cassava mash samples. Five grams of each sample was homogenized in 50 mL of distilled water and filtered. One milliliter of each filtrate and standard cyanide solutions [0.1 to 3 ppm KCN in distilled water and 20% HCl (1:1)] was measured into respective test tubes. 4 mL of alkaline picrate solution (1 g of picrate and 5 g of Na2CO3 in 200 mL of distilled water) was added to each test tube and incubated in a water bath (50°C/5 min) for development of dark-red color. The absorbance was taken at 490 nm (Halake et al., 2019).
The replicate values were reported as means±SD. Data were first subjected to one-way ANOVA, and then further analyzed using Tukey’s Honestly Significant Difference (HSD) post-hoc test. T-test was performed to confirm significant difference between the mean value of the uninoculated (control) group and inoculated group in the cassava mash fermentation studies. A p-value below 0.05 was defined as the criterion for statistical significance.
3. Results and discussion
All 17 samples of fufu and gari analyzed in this study were acidic (pH 3.13-4.78) and revealed occurrence of yeasts after cultivation on PDA, MEA and YPDA, where yeast colonies appeared whitish or creamy, smooth or rough, raised or flat and entire or undulating. Viable counts of yeasts were within the range of 4.00 and 6.64 Log CFU/mL (Supplementary Table S1). A total of 169 yeast isolates were randomly recovered from the three different isolation media, including 70 (41.42%), 52 (30.77%) and 47 (27.81%) isolates obtained from MEA, PDA and YPDA respectively. Exactly 61 (36.10%) and 108 (63.90%) yeast isolates were respectively recovered from nine samples of fufu (submerged fermentation) and eight samples of gari (solid state fermentation). Yeast isolates demonstrated diversity in cell shape and sugar fermentation pattern. The cells were either spherical, oval or rod shaped and they all fermented glucose, fructose, maltose and sucrose. They all grew to different extent at 1% acetic acid and produced acid from glucose.
Yeasts are an important group of microorganisms involved in the fermentation of cassava (Johansen et al., 2019; Oyewole, 2001). This study further confirms the predominance of wide diversity of yeasts during submerged and solid-state fermentation of cassava to produce fufu and gari, respectively. In a previous study, high density of up to 6.25 log CFU/g of yeasts were enumerated and identified in attiéké, a fermented cassava food in Burkina faso (Flibert et al., 2021).
Twenty (11.83%) of the yeast isolates showed potentials for protease production at different levels, while three (1.78%) isolates demonstrated amylase production potentials (Fig. 1). One hundred and fifty-seven yeast (92.31%) isolates released p-nitrophenol from p-nitrophenyl β-D-glucosidase (PNPG), indicating production of β-glucosidase to different extent, including high (54 isolates), moderate (47 isolates) and low (56 isolates) producers (Fig. 2). Three isolates that produced the three enzymes were selected and two isolates with high β-glucosidase production were randomly selected for further studies. They include isolates MOGB53, YOGB61, YOGB62, 3MOFA33 and 3YOGA45.
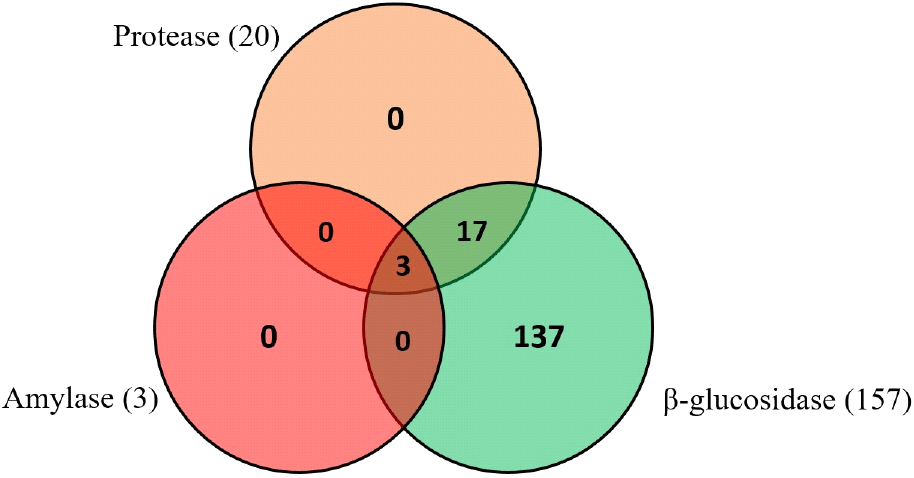
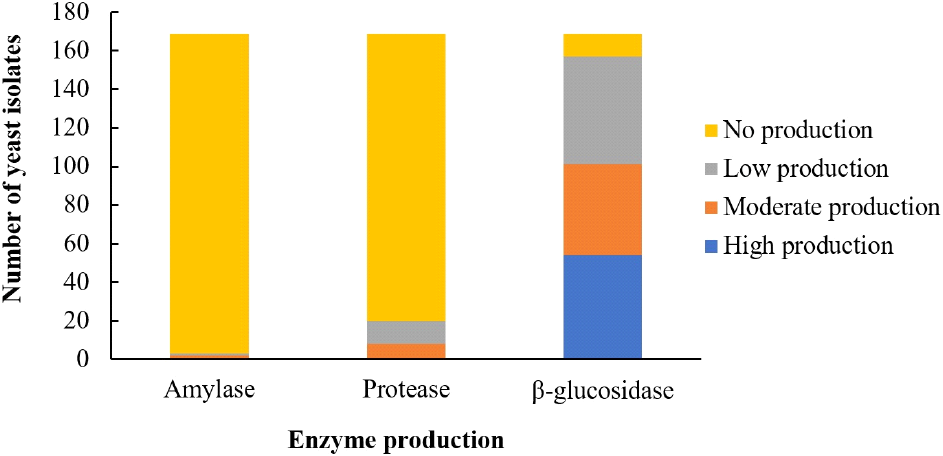
Enzymes produced by yeasts in cassava fermentation convert complex cassava components to simpler forms, thereby imparting desirable qualities on the products. This study revealed the limited distribution of amylase and protease production among yeast isolates from fufu and gari, while β-glucosidase production was widely distributed. The production of these enzymes was at different levels, varying from low, moderate to high producers. Amylase production by yeast isolated from gari and fufu was reported in previous studies (Aghemwenhio et al., 2017; Edward et al., 2012; Oyewole, 2001). Similarly, the potentials of yeast to secrete protease into food substrate was reported by Ogunremi et al. (2015) and (Aghemwenhio et al., 2017). Yeast amylase may breakdown cassava starch to simple sugar, thereby increasing the energy density of cassava and providing carbon source for LAB (Ogunremi and Sanni, 2011). Protease hydrolyzes proteins to amino acids and peptides. These metabolites act as flavour and/or bioactive compounds, thereby imparting the appeal and health benefits of fermented cassava foods. The demonstration of β-glucosidase production potential by most of the yeast isolates obtained in this study indicates their potential contribution to degradation of cyanogenic glucosides during cassava fermentation (Obilie et al., 2004). Microbial production of β-glucosidase could be largely induced by availability of enzyme-specific substrate, being cyanogenic glucosides (Okon et al., 2024).
Yeast growth was relatively inhibited by respective stress factor. None of the treatments presented growth index that is ≥100%. Isolate 3YOGA45 demonstrated the highest growth (98.42%) in YPD broth supplemented with 800 mg/L of cyanide, while the lowest growth was recorded for isolate YOGB61 (34.92%). Increasing growth inhibition was observed with increasing concentration of lactic acid. The growth indices of the five selected isolates at 1.20% lactic acid supplementation ranged from 34.67% to 63.62% (Fig. 3).
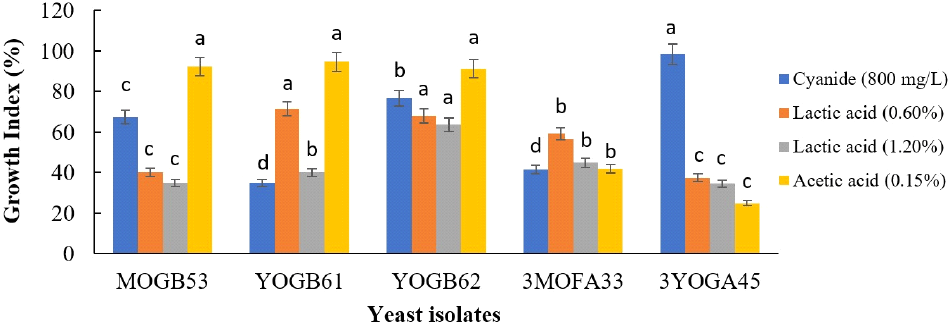
Yeast strains that are selected for controlled fermentation processes must tolerate typical stress conditions during the fermentation of specific substrate (Ogunremi et al., 2017). The presence of cyanide and accumulation of organic acids are potential stressors of microorganisms during the fermentation of cassava. Hydrogen cyanide inhibits the electron transport system in cells, thereby, disrupting respiration and energy metabolism. Organic acids cause acid stress and viability loss in microorganisms by diffusing across the membrane in protonated forms and subsequently dissociating intracellularly (Peyer et al., 2016). Cyanide levels ranging from 61.03 to 181.33 mg/kg were reported in various wild and improved cultivars of cassava (Njankouo Ndam et al., 2019). The average range of cyanide in fresh cassava root within sub-Saharan Africa is from ~100 ppm to ~450 ppm (Kobawila et al., 2005). In cases of high temperature, drought and CO2 pressure, cyanide level above 800 ppm has been reported (Cardoso et al., 2005; Nyaika et al., 2024).
In this study, growths were reported for the selected yeast isolates at high concentration of cyanide (800 mg/L), lactic acid (1.20%) and acetic acid (0.15%). In agreement with this study, Pichia exigua isolated from cassava mill effluent was reported to tolerate high concentration of cyanide (Banwo et al., 2023). The results from this study compare favourably with Ogbonnaya (2016), that reported increased growth of bacterial isolates, including Lactiplantibacillus plantarum, Limosilactobacillus fermentum, Lactobacillus amylovorus, Leuconostoc mesenteroides, Pseudomonas stutzeri, Bacillus pumilus and B. subtilis in medium containing 800 mg/L of cyanide. Tolerance to organic acids is well distributed among yeasts isolated from different food substrates (Bonatsou et al., 2018; Perricone et al., 2014).
Five selected yeast isolates that are technologically relevant for cassava fermentation, including MOGB53, YOGB61, YOGB62, 3MOFA33 and 3YOGA45 significantly survived in YPD broth at pH 2.5 and pH 3.5 and 0.3% and 0.5% bile salts (Table 1). The survival of the selected yeast isolates at pH 3.5 ranged from 62.50% to 92.71%. They were more sensitive at pH 2.5. The yeast isolates survived in YPD broth supplemented with bile salts. Their survival decreased as the concentration increased from 0.3% and 0.5% bile salts. Isolate YOGB62 demonstrated the highest survival at pHs 3.5 and 2.5 and 0.3% and 0.5% bile salts concentrations. The yeast isolates survived in simulated gastric juice (68.01-91.57%) and intestinal juice (54.16-87.58%).
A candidate microorganism for selection as potential probiotics must survive the harsh conditions in the gastrointestinal tract, including low pH, high concentration of bile salts, presence of hydrolytic enzymes and antimicrobial peptides (Staniszewski and Kordowska-Wiater, 2021). The pH in the stomach and bile salts concentration in the intestine at fed state are lethal to most microbes (Czerucka et al., 2007). This study is one of the few efforts to reports the survival of β-glucosidase-producing yeast strains isolated from cassava-based fermented foods at pH 3.5 and 2.5, 0.3% and 0.5% bile salts concentrations and simulated conditions in the stomach and intestine. This demonstrates that they are candidate strains for the detoxification of cassava and delivery of β-glucosidase in the gastrointestinal tract.
The adherence of selected yeasts isolates to two hydrocarbons are presented in Fig. 4. Isolate YOGB62 showed the highest adherence to toluene and xylene at 36.35% and 25% respectively. The lowest adherence to toluene was shown by isolate 3MOFA33 (3.54%), while it was shown by isolate MOGB53 (1.35%) to xylene. The percentage autoaggregation by the yeast isolates increased over a period of 24 h, ranging from 89.10% to 99.60% at 24h (Fig. 5).
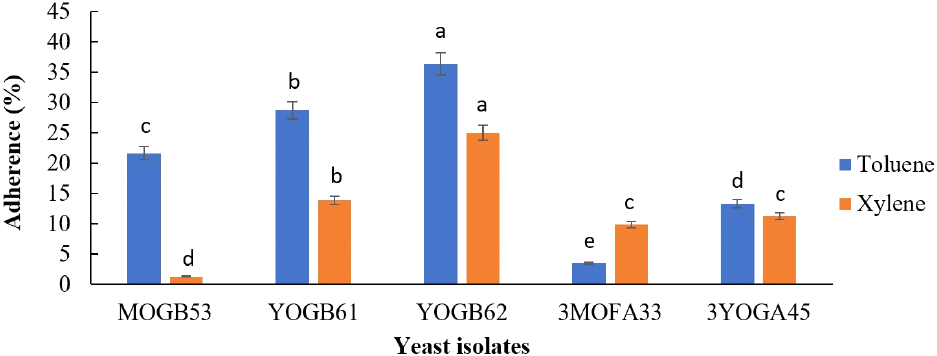
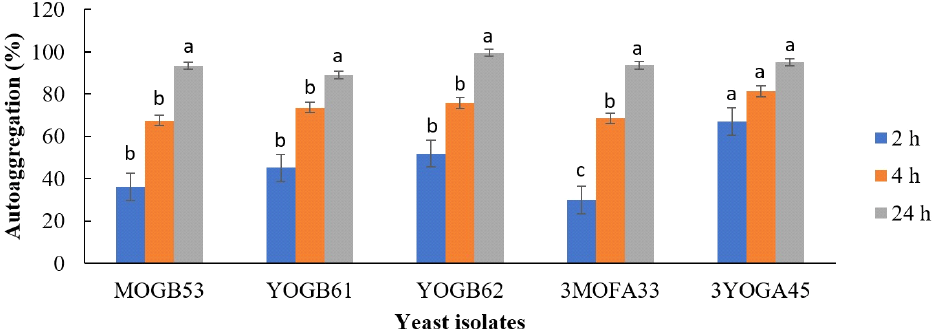
Besides regular consumption, the ability of such strains to persist in the intestine is important for stable supply of β-glucosidase and other bioactive compounds in the gut. This attribute is demonstrated by ability of probiotic strains to adhere to the surface of the epithelial cells and mucus and may be predicted as capacities to adhere to hydrocarbons and autoaggregate (Staniszewski and Kordowska-Wiater, 2021). The selected yeast strains in this study recorded hydrophobicity to toluene and xylene. Isolate YOGB61 and YOGB62 showed relatively stronger hydrophobicity to both hydrocarbons, ranging between 14 and 36%. In agreement with this study, Akinyemi et al. (2024) reported hydrophobicity to toluene and xylene falling within similar range for two strains each of Candida tropicalis and Diutina rugosa. Higher hydrophobicities (15-85%) to toluene and xylene were recently reported for four Pichia kudriavzevii strains isolated from ogi, a cereal-based weaning gruel in Nigeria (Ezekiel et al., 2025). Autoaggregation values above 40% is considered favourable for selection of potential probiotics, thereby providing possible colonization advantage in the gut (Akinyemi et al., 2024; García-Cayuela et al., 2014). The selected yeast isolates in this study showed autoaggregation percentage above 90%. Similarly, high autoaggregations have been reported for different species and strains of yeasts (Bonatsou et al., 2018; Ezekiel et al., 2025). The relatively bigger size and heavier weight of yeasts, compared to bacteria is expected to influence rapid and efficient precipitation (Gil-Rodríguez et al., 2015). The selected yeast isolates in this study possess the essential traits associated with potential probiotics, including potentials to survive the harsh conditions during gastrointestinal transit and persist in the gut.
The intracellular cell free extract of the yeast isolates scavenged 200 μM DPPH free radicals in methanol reaction system at different levels (Table 2). DPPH scavenging activity of the yeast isolates ranged from 4.00% to 19.74%, with isolate MOGB53 demonstrating the strongest DPPH scavenging activity.
| Yeast isolate | DPPH scavenging activity (%) |
|---|---|
| MOGB53 | 19.74±1.731)a2) |
| YOGB61 | 8.06±0.29b |
| YOGB62 | 4.00±0.08c |
| 3MOFA33 | 4.97±0.68c |
| 3YOGA45 | 4.00±0.20c |
In addition, to their potentials to deliver β-glucosidase, they may also support the scavenging of reactive oxygen species in the host. Antioxidants prevent the oxidation of important biomolecules, such as nucleic acids, proteins and lipids, thereby, reducing the risk of degenerative diseases (Bloomer et al., 2011). The antioxidative potentials of yeasts have been richly reported (Akinyemi et al., 2024; Ezekiel et al., 2025; Ogunremi et al., 2015).
Yeast isolate “YOGB62” was selected based its versatile enzyme profile, survival at in-vitro stress factor during cassava fermentation and probiotic potentials, particularly, high β-glucosidase production and high survival at high cyanide concentration, low pH, different concentrations of bile salts and simulated gastrointestinal conditions. The yeast isolate was identified by gene sequence analysis on NCBI database (NCBI, 2025) as Wickerhamomyces anomalus YOGB62 (Accession number: OP048088).
The yeast strain with relatively greater prospects of β-glucosidase production, stress tolerance and probiotic attributes that was selected and genetically identified as Wickerhamomyces anomalus YOGB62 is of prominent biotechnological interest. It is listed in the category of microorganisms for beneficial technological importance (Bourdichon et al., 2012). W. anomalus was reported to be predominant among the yeast species isolated from artisan bakery sourdoughs and spontaneous laboratory sourdoughs (Vrancken et al., 2010). W. anomalus was also the most dominant yeast species at different stages of cassava processing to produce rale, a cassava roasted flour in Mozambique (Massamby et al., 2025).
Cassava mash supported the growth of W. anomalus YOGB62, with cell density increasing by over 2.0 log units after 72 h of fermentation (Fig. 6A). The controlled fermentation with W. anomalus YOGB62 significantly acidified the cassava mash, with reduction of the pH from 7.26 to 3.39 after 72 h (Fig. 6B). The cyanide degradation potential of W. anomalus YOGB62 was demonstrated as presented in Fig. 6C. Controlled fermentation of cassava mash with W. anomalus YOGB62 resulted to reduction of cyanide content from 40.15 mg/kg to 10.82 mg/kg and an undetectable level after 48 h and 72 h of fermentation, respectively (Fig. 6C). This is contrary to what applies to the control sample with significant proportion of cyanide still retained after 72 h.
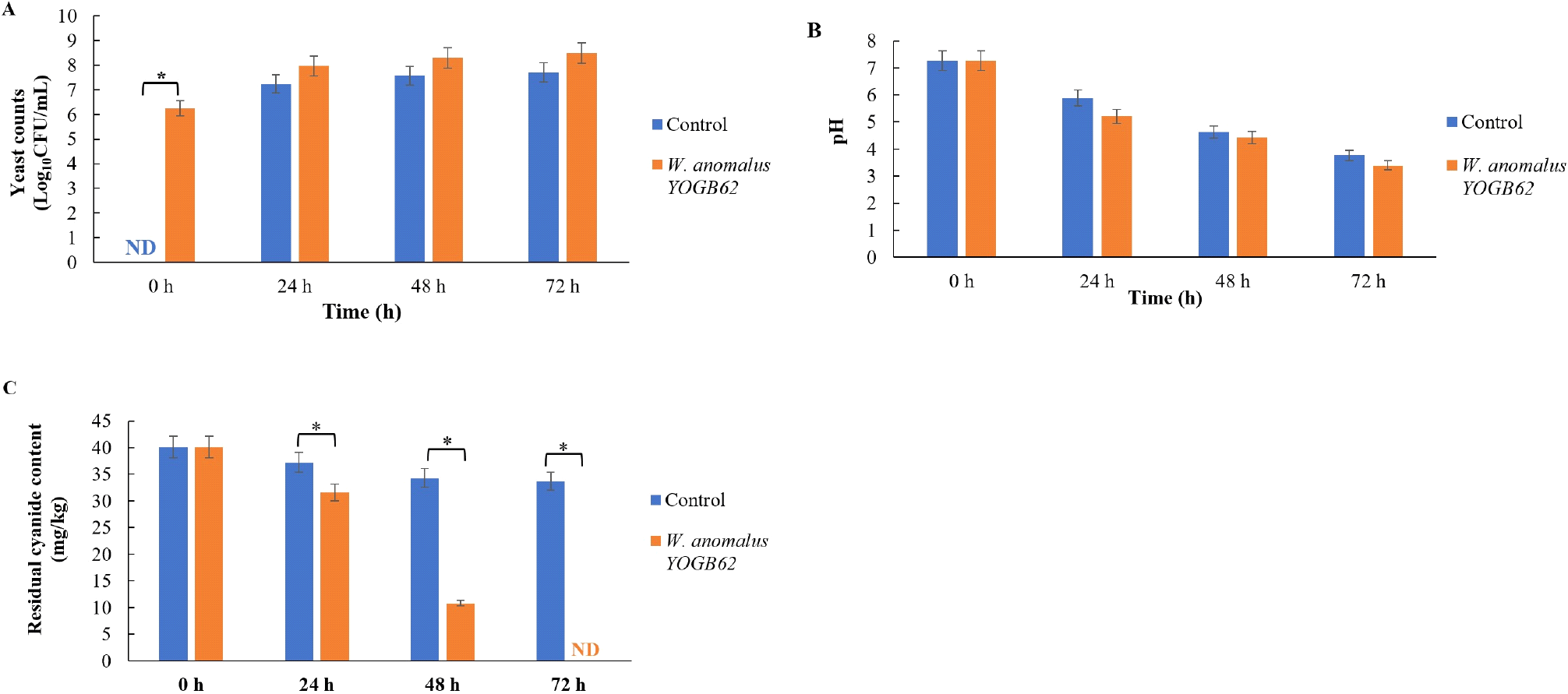
In this study, the growth of W. anomalus YOGB62 inoculum in cassava mash by approximately 2 log units, coupled with reduction in pH are indications of the suitability of the strain to metabolise components of cassava mash. In agreement with this study, a previous study reported that Saccharomyces cerevisiae inoculum reduced the pH of cassava flour (Halake et al., 2019). The robust elaboration of β-glucosidase from W. anomalus YOGB62 as a single-strain inoculum in the fermentation of cassava mash must be responsible for the degradation of cyanide to an undetectable level after 72 h. Similar, excellent degradation of cyanide by single-strain starter cultures of systematically selected yeasts have been demonstrated during controlled fermentation of cassava substrate (Banwo et al., 2023; Halake et al., 2019).
4. Conclusions
This study revealed the predominant occurrence of β-glucosidase-producing and potentially probiotic yeasts in fufu and gari. The selected yeast demonstrated potentials for application as probiotic starter cultures to detoxify cassava-based fermented foods and derive potentially healthful products. This may constitute an affordable and sustainable strategy to optimally utilize cassava.