
1. Introduction
Skin aging is not merely a result of excessive reactive oxygen species (ROS) production but is also driven by oxidative imbalance arising from intrinsic factors such as cellular senescence and hormonal changes, as well as extrinsic stimuli including ultraviolet (UV) radiation, environmental pollution, and smoking (Fisher et al., 2002; Liang et al., 2023). These factors collectively contribute to increased generation of ROS, disrupting redox homeostasis in skin tissue (Rinnerthaler et al., 2015). In particular, UV radiation increases the expression of matrix metalloproteinases (MMPs) and elastase, promoting collagen degradation and reducing skin elasticity (Poljšak and Dahmane, 2012). Therefore, inhibition of ROS generation and reinforcement of endogenous antioxidant systems are widely considered essential strategies for mitigating skin aging. Natural antioxidant compounds such as polyphenols, flavonoids, and vitamins derived from plant sources have received considerable attention as functional ingredients for skin protection due to their free radical scavenging, anti-inflammatory, and collagen-related activities (Tomas et al., 2025). These compounds ameliorate oxidative stress through coordinated regulation of redox-sensitive pathways, neutralize ROS, and modulate endogenous antioxidant enzymes including superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx), thereby limiting oxidative damage in skin tissue (Rinnerthaler et al., 2015).
Hemerocallis fulva L. (daylily) is a perennial plant belonging to the family Liliaceae and has been traditionally utilized as both a food material and medicinal resource in Korea, China, and Japan (Hsu et al., 2023; Wang et al., 2022). The plant is rich in amino acids, organic acids, and β-carotene and has demonstrated a broad spectrum of biological activities, including antioxidant, anti-inflammatory, hepatoprotective, and antidepressant effects (Fu and Mao, 2008; Hsu et al., 2023; Liu et al., 2022; Yan et al., 2023). These functional effects are primarily attributed to phenolic acids and flavonoid compounds that contribute to redox regulation via electron-donating capacity and metal ion chelation mechanisms (Szewczyk et al., 2020). However, previous studies on H. fulva have mainly focused on its flowers and roots, with little research on the biological activities and functional properties of the edible leaves. Given their safety and applicability as dietary resources, leaves represent a promising material for functional development, and evaluation of solvent-dependent extraction efficiency is therefore of practical significance for industrial applications. Accordingly, this study aimed to evaluate the antioxidant activities (DPPH, ABTS, FRAP, and SOD-like assays) and wrinkle-related functional markers (elastase inhibition and stimulation of type I collagen synthesis) of water extract (HF-WE) and ethanol extract (HF-EE) from young leaves of H. fulva using an in vitro screening approach.
2. Materials and methods
Young leaves of H. fulva were collected during the initial vegetative phase (March-April 2022) from Yecheon, Gyeongbuk, Korea. The leaves were washed thoroughly with water, air-equilibrated at ambient temperature (25°C) to remove surface moisture, and subsequently freeze-dried. The dried materials were stored under frozen conditions (-20°C) prior to further experimental use.
All solvents used in this study were of analytical grade and were purchased from Duksan Pure Chemicals Co., Ltd. (Ansan, Korea). DPPH (2,2-diphenyl-1-picrylhydrazyl), ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)), Folin-Ciocalteu reagent, rutin, quercetin, and L-ascorbic acid were procured from commercial sources (Sigma-Aldrich, St. Louis, MO, USA). Fetal bovine serum (FBS) was provided by Alphabioregen (Burlington, MA, USA). High-glucose Dulbecco’s modified Eagle’s medium (DMEM), L-glutamine, sodium pyruvate, and penicillin-streptomycin were acquired from HyClone (Waltham, MA, USA). Tetrazolium bromide (MTT) and dimethyl sulfoxide (DMSO) were also obtained from Sigma-Aldrich. Elastatinal (Sigma-Aldrich) was employed as a reference inhibitor in the elastase assay. The Procollagen Type I C-peptide (PIP) EIA kit was supplied by Takara (Otsu, Japan).
Freeze-dried H. fulva leaf powder (10 g) was extracted in independent batches (n=3) with either distilled water or 70% ethanol at a solvent-to-sample ratio of 10:1 (mL/g). Each extraction was carried out for 5 h under controlled conditions (25°C) with gentle agitation. The resulting extracts were filtered through Advantec No. 2 qualitative paper (Toyo Roshi Kaisha Ltd., Tokyo, Japan), and the pooled filtrates were reduced in volume using rotary evaporation under reduced pressure at temperatures not exceeding 40°C (N-1000, EYELA, Tokyo, Japan). Final concentrates were freeze-dried to constant mass (7 days) and kept at -20°C pending analysis. The aqueous and hydroethanolic preparations were hereafter referred to as HF-WE and HF-EE, respectively.
Total polyphenol content was determined by a Folin-Ciocalteu colorimetric assay. Extract (1 mL) was mixed with distilled water (5 mL) and Folin-Ciocalteu reagent (0.5 mL), followed by addition of 7% Na2CO3 (10 mL). After adjustment to 25 mL, the mixture was incubated in the dark for 2 h, and absorbance was measured at 750 nm (UV-1800, Shimadzu, Kyoto, Japan). Results were expressed as mg gallic acid equivalents (GAE) per gram of dry weight (DW) of sample. Total flavonoid content was measured using an AlCl3 colorimetric method. Extract (40 μL) was reacted sequentially with 5% NaNO2 (6 μL), 10% AlCl3 ․ 6H2O (12 μL), and 1 N NaOH (500 μL). Absorbance was recorded at 510 nm, and data were expressed as mg rutin equivalent (RE) per gram of DW of sample.
Extract (0.2 mL) was mixed with 0.2 mM DPPH solution in ethanol (0.8 mL), incubated in the dark for 30 min, and absorbance was measured at 517 nm. Radical scavenging activity was expressed as percentage inhibition.
ABTS•+ radicals were produced by reacting ABTS (7.4 mM) with potassium persulfate (2.6 mM) for 18 h at 25°C in the dark. The solution was diluted to an absorbance of 0.70 at 734 nm before analysis. Sample (40 μL) was reacted with ABTS•+ solution (4.0 mL) for 1 min, and absorbance was measured at 734 nm. Results were calculated as percentage inhibition using L-ascorbic acid as reference.
FRAP reagent was freshly prepared by mixing acetate buffer (pH 3.6), TPTZ, and FeCl3 ․ 6H2O at a ratio of 10:1:1 (v/v/v). Sample (0.5 mL) was reacted with FRAP reagent (3.5 mL), and absorbance at 593 nm was recorded after 5 min at 37°C. Results were expressed as μM FeSO4 equivalents/g sample.
SOD-like activity was evaluated using a pyrogallol autoxidation-based inhibition assay adapted from Marklund and Marklund (1974). Aliquots of extracts (10 μL) were added to individual wells of a microplate, followed by Tris-HCl buffer (50 mM Tris, 10 mM EDTA, pH 8.5; 150 μL) and pyrogallol solution (7.2 mM; 50 μL). The reaction proceeded at 25°C for 45 min and was halted by adding HCl (1 N; 50 μL). Absorbance was read at 420 nm using a microplate-based optical system, and activity was expressed as inhibition percentage of pyrogallol oxidation.
Elastase inhibition was assessed by monitoring substrate cleavage under controlled conditions following Bieth et al. (1974) with minor modification. The reaction system contained N-succinyl-Ala-Ala-Ala-p-nitroanilide (1.0 mM in 0.1 M Tris-Cl, pH 8.0; 1,300 μL), sample (7.5 μL), and buffer (92.5 μL). After equilibration at 25°C for 10 min, elastase solution (10 μg/mL in 0.12 M Tris-Cl; final 0.0025 U/mL; 100 μL) was added to trigger the reaction. Following incubation at 25°C for 20 min, absorbance at 410 nm was recorded, and inhibition was calculated as a relative decrease versus the control.
Cytotoxicity toward CCD-986Sk fibroblasts was examined by quantifying metabolic activity using the MTT assay. Cells were seeded in standard culture plates (1.3×105 cells/mL) and allowed to adhere for 24 h prior to treatment. After 24 h exposure to extracts, MTT solution (200 μg/mL) was added and incubation continued for 3 h at 37°C. Formazan was dissolved in DMSO (100 μL), absorbance was measured at 570 nm, and viability was reported as percentage relative to untreated cells.
After 24 h treatment, culture supernatants were collected for analysis of type I procollagen using a commercial enzyme immunoassay (PIP EIA kit, Takara). Assays were conducted according to the supplier’s procedure; reactions were developed at 25°C for 15 min and stopped with H2SO4 (1 N; 100 μL). Absorbance at 450 nm was measured, and concentrations were derived from the assay-specific calibration curve.
Data are presented as mean±SD, and analyses were performed using SPSS (ver. 25). For comparisons involving multiple groups, one-way analysis of variance (ANOVA) was applied, followed by Duncan’s multiple range test. Student’s t-test was used to determine significant differences between two extract groups (HF-WE and HF-EE). Differences were considered statistically significant at p<0.05.
3. Results and discussion
The extraction yields of HF-WE and HF-EE were 17.47% and 20.35%, respectively. Based on these extracts, the total polyphenol contents were determined, and HF-EE exhibited a significantly higher polyphenol content (27.26 mg GAE/g DW) than HF-WE (18.52 mg GAE/g DW) (p<0.001) (Table 1). The total flavonoid contents were 8.93 mg RE/g DW for HF-WE and 12.44 mg RE/g DW for HF-EE, demonstrating a significantly greater accumulation of flavonoids in the ethanol extract (p<0.001). These results are in good agreement with previous reports by Thiruvengadam et al. (2014) and Byun et al. (2021), which showed that ethanol extraction results in higher recovery of phenolic compounds than hot-water extraction in leafy plant resources, including Aster scaber and Sanguisorba officinalis. This tendency may be explained by the physicochemical characteristics of ethanol, which functions as a medium-polarity solvent capable of dissolving both hydrophilic and hydrophobic constituents. Because polyphenolic molecules contain both hydrophilic hydroxyl groups and hydrophobic aromatic structures, ethanol enables more effective extraction efficiency compared to water (Alara et al., 2021; Wang et al., 2022).
| Sample1) | Total polyphenol contents (mg GAE/g DW) | Total flavonoid contents (mg RE/g DW) |
|---|---|---|
| HF-WE | 18.52±0.15 | 8.93±0.04 |
| HF-EE | 27.26±0.032)***3) | 12.44±0.57*** |
The DPPH radical scavenging activities of HF-WE and HF-EE are shown in Fig. 1A. Both extracts demonstrated concentration-dependent increases in scavenging activity, with HF-EE consistently outperforming HF-WE across the tested concentration range. Specifically, the scavenging activity of HF-WE increased from 4.33% to 70.35% at concentrations of 50-800 μg/mL, whereas HF-EE increased from 4.51% to 87.20% under the same conditions, indicating a significantly greater antioxidant capacity than HF-WE (p<0.05). The DPPH method is based on the reduction of the stable free radical by electron or hydrogen donation from antioxidant molecules, which leads to a measurable decline in absorbance (Blois, 1958). The superior scavenging activity of HF-EE can be reasonably associated with its higher polyphenol and flavonoid contents, as these compounds possess hydroxyl groups capable of stabilizing free radicals through electron donation mechanisms (Dai and Mumper, 2010; Lohvina et al., 2022). A similar pattern has been documented in S. officinalis extracts, where ethanol extraction resulted in greater polyphenolic yield and enhanced DPPH radical scavenging effects compared with water extraction (Byun et al., 2021), which is consistent with the trend observed in the present study.
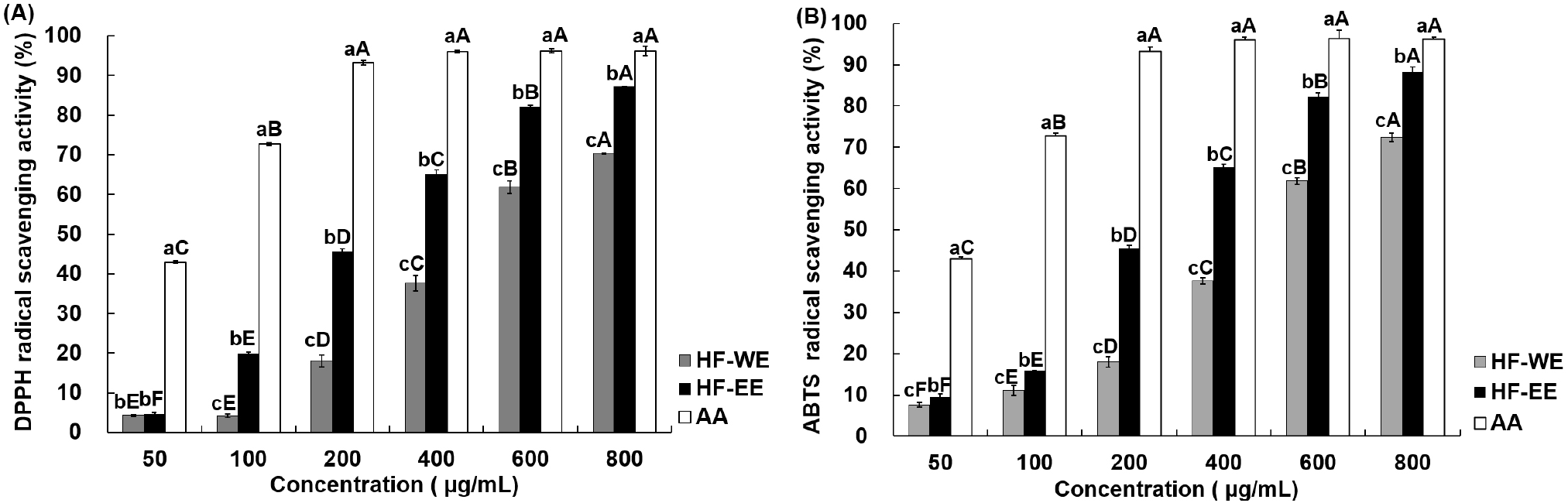
The ABTS radical scavenging activities of HF-WE and HF-EE are shown in Fig. 1B. The scavenging activity of HF-WE increased from 7.60% at 50 μg/mL to 72.40% at 800 μg/mL, whereas that of HF-EE rose from 9.51% to 88.23% within the same concentration range. Both extracts exhibited concentration-dependent responses, and HF-EE showed a significantly higher ABTS radical scavenging activity than HF-WE at all tested concentrations (p<0.05). The ABTS method involves the reduction of the ABTS•+ cation radical by electron-donating antioxidants, resulting in discoloration of the radical chromophore and allowing indirect evaluation of electron transfer capacity (Re et al., 1999). The stronger radical scavenging ability of HF-EE can be attributed to its higher content of phenolic and flavonoid constituents, which are widely recognized to donate electrons and stabilize free radicals (Dai and Mumper, 2010; Szewczyk et al., 2020). Although HF-WE displayed lower absolute activity values, its dose-dependent increase demonstrates that H. fulva leaves exhibit substantial antioxidant capacity regardless of the extraction solvent.
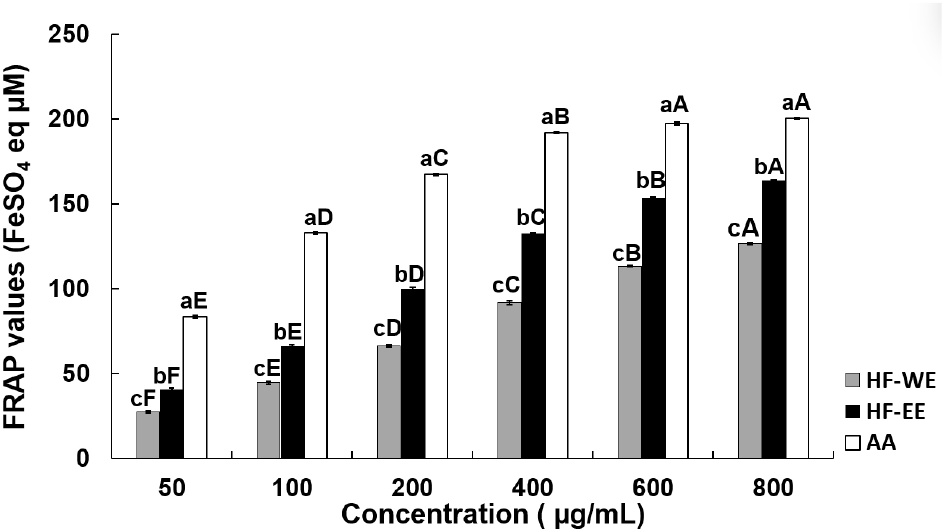
The ferric reducing antioxidant power of HF-WE and HF-EE is shown in Fig. 2. Both extracts displayed significant, concentration-dependent increases in FRAP values (p<0.05). The FRAP value of HF-WE increased from 27.41 μM FeSO4 equivalents/g at 50 μg/mL to 126.63 μM FeSO4 equivalents/g at 800 μg/mL, whereas HF-EE exhibited higher reducing capacity, ranging from 40.63 to 164.08 μM FeSO4 equivalents under identical concentration conditions (p<0.05). The FRAP assay evaluates antioxidant reducing power by quantifying the conversion of ferric ions (Fe3+) into ferrous ions (Fe2+), and elevated values correspond to increased reducing potential (Benzie and Strain, 1996). Polyphenolic compounds such as quercetin, rutin, and caffeic acid are widely recognized as strong reducing agents due to their electron-rich molecular structures. Accordingly, the superior FRAP activity of HF-EE is likely explained by its higher phenolic and flavonoid contents. In support of this interpretation, Szewczyk et al. (2020) reported that aerial extracts of the Hemerocallis genus contain abundant quercetin derivatives, including quercetin-3-O-rutinoside. Collectively, these findings suggest that ethanol extraction improves the recovery of phenolic antioxidants, thereby contributing to the superior reducing power observed in HF-EE.
The SOD-like activities of HF-WE and HF-EE are shown in Table 2. Both samples exhibited concentration-dependent increases in SOD-like activity with statistically significant differences (p<0.05). HF-EE exhibited significantly greater activity than HF-WE at all tested concentrations except 50 μg/mL (p<0.05). In addition, the antioxidant effect of HF-EE was comparable to that of Trolox at selected concentrations (5, 200, and 500 μg/mL), supporting its strong superoxide-scavenging capacity. SOD-like activity reflects the ability of antioxidant compounds to reduce superoxide radical formation through redox-related mechanisms rather than a single removal pathway. These effects have been commonly linked to the presence of phenolic and flavonoid compounds (Esmaeili et al., 2015). H. fulva is known to contain high levels of phenolic constituents, including chlorogenic acid, rutin, and quercetin derivatives (Fu and Mao, 2008; Hao et al., 2022), which are reported to participate in antioxidant defense via mechanisms such as electron transfer, metal ion binding, and radical quenching. Accordingly, the higher SOD-like activity observed in HF-EE is likely related to its comparatively greater enrichment of redox-active phenolic compounds relative to HF-WE.
HF-WE, water extract of H. fulva; HF-EE, ethanol extract of H. fulva; Trolox, 6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid, positive control.
Different lowercase letters (a-c) within the same column indicate significant differences among samples at the same concentration (p<0.05). Different uppercase letters (A-F) within the same row indicate significant differences among concentrations within the same sample (p<0.05), based on Duncan’s multiple range test.
The elastase inhibitory activities of HF-WE and HF-EE are shown in Table 3. Both extracts showed a dose-responsive increase in elastase inhibition with statistical significance (p<0.05). At concentrations of 200 μg/mL and above, inhibition values greater than 80% were observed in both samples, indicating substantial inhibitory activity. No meaningful difference was detected between HF-WE and HF-EE across the tested concentration range, and both HF-WE and HF-EE reached approximately 88% inhibition at 500 μg/mL, which was comparable to that of the positive control, elastatinal. Elastase is a proteolytic enzyme that degrades elastin, a major structural protein of the dermal extracellular matrix, and excessive elastase activity is known to promote skin aging by causing structural damage to elastin fibers (Schmelzer et al., 2012). In the present study, HF-EE did not exhibit a proportional relationship between total polyphenol and total flavonoid contents and elastase inhibitory activity when compared with HF-WE. Previous studies have reported elastase inhibitory activity in plant extracts with high polyphenol and flavonoid contents, suggesting that these compounds may contribute to elastase inhibition (Radjah et al., 2021). In addition, certain natural flavonoids have been reported to inhibit elastase activity through direct interactions with the enzyme active site (Lin et al., 2025). In contrast, other studies have demonstrated that elastase inhibitory activity does not consistently show a linear relationship with total polyphenol content or antioxidant capacity, and that various bioactive constituents, including water-soluble polysaccharides, glycosylated phenolic compounds, and other non-phenolic components, may influence elastase inhibition (Eun et al., 2020). Taken together, the elastase inhibitory activities observed for HF-WE and HF-EE in the present study can be interpreted as in vitro experimental outcomes that are influenced by phenolic and flavonoid compounds, but are likely regulated by the compositional complexity and interactions among diverse bioactive constituents present in the extracts. Therefore, further studies are warranted to identify the key active compounds responsible for elastase inhibition and to elucidate their underlying mechanisms of action.
HF-WE, water extract of H. fulva; HF-EE, ethanol extract of H. fulva; Elastatinal, positive control.
Different lowercase letters (a-c) within the same column indicate significant differences among samples at the same concentration (p<0.05). Different uppercase letters (A-F) within the same row indicate significant differences among concentrations within the same sample (p<0.05), based on Duncan’s multiple range test.
The cytotoxic effects of HF-WE and HF-EE in CCD-986Sk cells are shown in Fig. 3A. Both extracts exhibited concentration-dependent reductions in cell viability within the range of 5-200 μg/mL (p<0.05). The viability of HF-WE decreased from 99.1% at 5 μg/mL to 78.8% at 200 μg/mL, whereas that of HF-EE was reduced from 98.6% to 74.0% over the same concentration interval. Since cell viability maintained above 70% across the tested concentrations, no marked cytotoxic effects were observed under these experimental conditions, making them suitable for subsequent functional evaluation (ISO 10993-5:2009). Based on these observations, concentration ranges that did not elicit appreciable cytotoxicity in the MTT assay were applied in subsequent experiments to evaluate type I collagen synthesis.
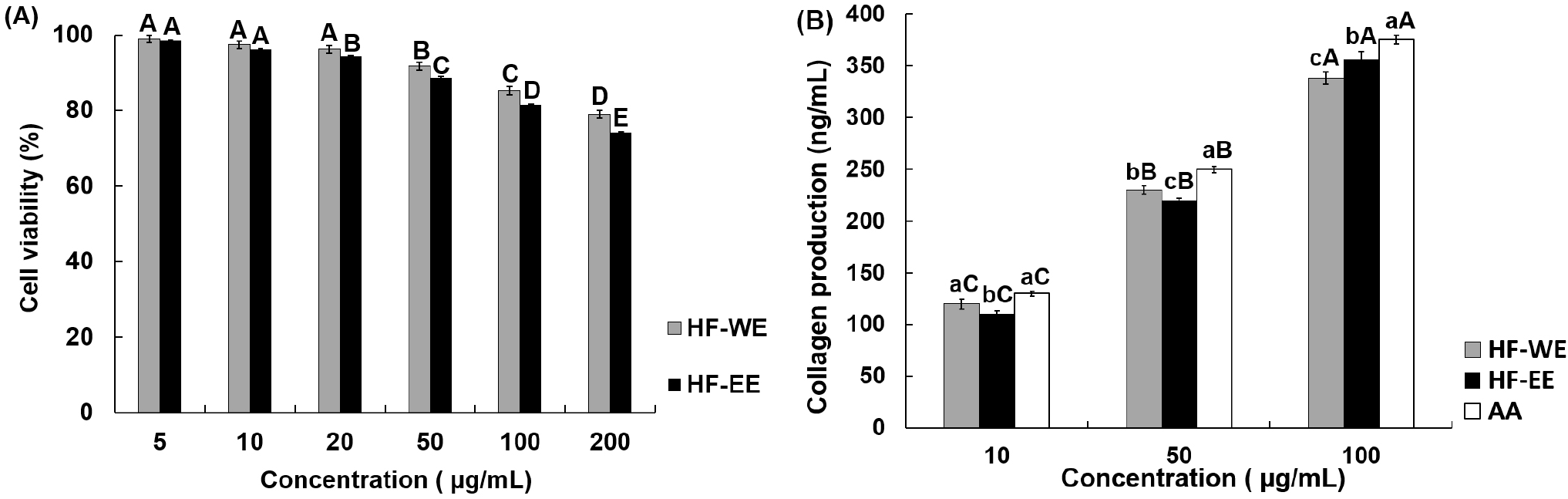
The effects of HF-WE and HF-EE on type I collagen synthesis in CCD-986Sk cells are shown in Fig. 3B. Both extracts demonstrated statistically significant, concentration-dependent elevations in type I collagen production (p<0.05). At 50 μg/mL, HF-WE yielded slightly greater collagen synthesis than HF-EE, whereas at concentrations of 100 μg/mL and above, collagen output induced by both extracts approached that of the positive control (ascorbic acid). The present findings suggest that H. fulva leaf extracts may enhance collagen production in human dermal fibroblasts under the experimental conditions employed. However, the underlying mechanisms responsible for this response cannot be directly determined based on the present data. Previous studies have reported that phenolic compounds contribute to the maintenance of collagen homeostasis by attenuating oxidative stress and suppressing matrix metalloproteinase (MMP)-mediated collagen degradation, thereby supporting extracellular matrix stability (Bjørklund et al., 2022). In light of these reports, the increased collagen production observed in the present study may be associated with antioxidant-related activity. Nevertheless, because mechanistic analyses were not conducted, this interpretation requires further verification in future studies.
4. Conclusions
In this study, HF-WE and HF-EE were comparatively evaluated in terms of antioxidant activity, bioactive compound content, and cellular responses under in vitro conditions. HF-EE exhibited significantly higher total polyphenol and flavonoid contents than HF-WE and showed overall stronger antioxidant activity in DPPH, ABTS, FRAP, and SOD-like assays. Both extracts did not induce marked cytotoxicity in human dermal fibroblasts and demonstrated significant elastase inhibitory activity and increased type I collagen synthesis within the tested concentration range. Overall, these results indicate that H. fulva leaf extracts exhibit positive responses in antioxidant activity and cellular responses under in vitro conditions. However, as the present study is based on in vitro evaluation, further in vivo studies, including animal models, will be necessary to more clearly elucidate the physiological relevance and potential applicability of these findings.