
1. 서론
우리나라 고유의 술인 탁·약주를 빚는 데 사용하는 전통누룩은 자연계에서 유래한 곰팡이, 효모, 젖산균 등 다양한 종류의 발효 미생물이 분포하고 있다(Jung과 Chung, 2017). 이러한 미생물은 전분분해 활성, 알코올 생산, 유기산과 유리아미노산 및 향기성분 등의 차이를 나타낸다(Kim 등, 2009; Lee 등, 2015; Nam 등, 2015). 지역별로 다양한 특성이 있는 전통누룩은 탁·약주 등을 빚는 발효제로써 사용되고 있으며, 쌀, 보리와 같은 잡곡류 등을 파쇄하여 무증자 상태로 성형한 후, 자연 발효를 통해 만들어진다. 누룩 속의 미생물 분포와 성분 변화는 제조한 지역, 원료, 제조 방법 등 지리적·환경적 조건에 의하여 결정되며(Kim 등, 2014b), 원료 또한 다양하여 옥수수, 보리, 대두, 귀리, 조 및 쌀 등의 잡곡류로 만들어 탁·약주 제조에 사용되었다(Baek 등, 2010). 누룩에 분포하는 미생물 중에서 알코올과 향미 생성에 관여하는 효모는 대부분 GRAS(generally recognized as safe) 균주로서, Saccharomyces cerevisiae를 비롯하여 non-Saccharomyces 계열인 병원균 Candida albicans의 길항작용으로 의약산업에 응용할 수 있는 Pichia jadinii (Buerth 등, 2016; Song 등, 2013), 고알코올 농도에 내성이 있는 P. anomala(Tao 등, 2011), 고온에서 glucoamylase 효소활성 및 비성장속도가 우수한 Saccharomycopsis fibuligera (Choi 등, 2014) 등이 분리된 바 있지만, Saccharomyces cerevisiae 이외 산업적으로 상용화되지는 못하였다. 특히, 국내 대부분 양조장에서 탁·약주 제조용 효모는 와인 및 제빵용 효모(La Parisinne, LP 등)를 수입하여 사용하는 실정이다.
주류에 있어서 알코올 생산과 풍미에 관여하는 효모는 진핵생물로서 오래전부터 주류 및 제빵 등의 식품산업뿐만 아니라, 사료용 첨가물, 균체 생산 및 유지와 색소생산 등 biotechnology(BT) 산업의 핵심 소재로써 이용된다(Kim 등, 1999; Lee 등, 1997; Lee 등, 1999). 학문적인 분야에서 특정 대사산물의 생성, 분비와 발현 기작 해석 등의 분자생물학적 도구로서 폭넓게 이용되고 있다(Kim 등, 2004). 최근에는 항고혈압성 안지오텐신 전환효소 저해물질(Kim 등, 2004), 항치매성 β-secretase 저해물질(Lee 등, 2007)과 acetylcholinesterase 저해물질(Lee 등, 2008), 혈관신생 억제물질(Jeong 등, 2006), 미백성 tyrosinase 저해물질(Jang 등, 2012b), 혈전용해 활성물질(Jang 등, 2011) 및 정미성 핵산 물질(Lee 등, 2004) 등 건강 기능성 관련 연구가 활발히 이루어지고 있다.
따라서, 향미 증진 등 발효 특성이 우수한 효모 종균을 발굴하여 우리 술의 품질 향상과 종균 산업 발전에 이바지하고자 본 연구에서는 알코올 생산성이 뛰어난 효모를 선발하여 이들의 기능적 특성 등을 규명하고자 하였다.
2. 재료 및 방법
발효 특성이 우수한 토착 효모를 분리·선발하기 위하여 국내 4개 지역(부산, 서울, 전북, 경기)에서 제조한 전통누룩을 수집하였다. 누룩에 자생하는 효모를 순수분리하기 위하여 YPD(BD DIFCO, Mississauga, Ontario, Canada) 배지에 sodium propionate를 0.45% 첨가하여 곰팡이의 생육을 억제시켰다. 생리식염수에 현탁한 누룩 1 g을 YPD 고체배지에 100 μL 도말하고 25°C에서 24-48시간 배양하여 단일 colony를 분리하였다. 분리된 효모는 YPD 액체배지에 배양하여 이후 실험을 위하여 배양액에 40% 글리세롤을 첨가해 −80°C 냉동고에 보관하였다. 비교 균주로는 국내 종균 수입업체 삼광산업(주)(Seoul, Korea)에서 구입한 Saccharomyces cerevisiae (La Parisinne, Saint-Denis, France)를 사용하였다.
수집 전통누룩으로부터 분리한 효모 동정은 18S rDNA 유전자 염기서열로 분석하였다. 프라이머는 NS1(5’-GT AGTCATATGCTTGTCTC-3’)과 NS8(5’-TCCGCAGGT TCACCTACG GA-3’)을 사용하였고, PCR은 initial denaturation 5분, 그리고 94°C에서 45초간 denaturation, 55°C에서 60초간 annealing 및 72°C에서 60초간 extension cycle을 35회 수행하였다. PCR 산물의 염기서열은 ㈜제노텍(GenoTech Corp., Daejeon, Korea)에 의뢰하여 분석하였고(Kim 등, 2008), phylogenetic tree는 The National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov/)에서 제공하는 Advanced Blast Search 프로그램을 통하여 GenBank에 보고된 표준 균주와의 염기서열을 비교하여 계통분류학적 유연관계를 분석한 후, MEGA V6.0을 이용하여 Tamura-Nei distance model과 neighborjoining method에 따라 계통도를 작성하였다(Tamura 등, 2013).
분리 효모의 온도 및 pH별 생육 특성을 규명하기 위해 YPD 고체배지에서 생균수를 측정하였다. 생균수 측정은 십진 희석법을 이용하여 YPD 고체배지에 100 μL를 도말하고 30°C에서 48시간 배양한 후, 생성된 집락 수를 계수해 형성 단위(colony forming units, CFU/mL)로 나타내었다.
효모의 알코올 생성량을 조사하기 위해 분리된 효모를 20% glucose(Chung 등, 1990a; Rhee 등, 1997)가 포함된 YPD 액체배지에 각각 접종하여 25°C에서 5일간 배양한 후, 원심분리(10,000 ×g, 5 min)한 상등액 100 mL를 시험관에 넣고 소포제(규소수지) 2-3방울을 떨어뜨려 증류기(VAP200, Gerhadt, Konigs-winter, Germany)로 증류하였다. 회수한 증류액 80 mL에 증류수를 가하여 100 mL로 정용한 후, 밀도측정기(AT/DMA5000M, Anton Paar, TR, USA)를 사용하여 측정하였다.
α-Amylase 활성은 효모 배양액을 13,000 rpm에서 5분간 원심분리한 상등액을 사용하였다. 각각의 배양 상등액 20 μL에 멸균수 180 μL를 가한 후, 30 mM PBS (pH 6)에 녹인 starch (1 g/L) 200 μL를 섞어 30°C에서 20분 배양하였다. 각각의 배양 검체에 DNS regent 400 μL를 넣어 5분간 끓인 후, 실온에서 냉각하여 흡광도(BioTek®, Gen5™, Winooski, VT, USA) 550 nm에서 α-amylase 활성을 측정하였다. 1 unit(U)의 효소활성은 1분 동안 1 μM의 환원당을 생성하는 효소의 양으로 정의하였다. 한편, 정성적인 방법은 Park 등(2018)의 방법을 따랐다. LB 배지(1% tryptone, 0.5% yeast extract, 1% sodium chloide)에 1% soluble starch를 넣고 24시간 배양 후, 요오드 용액(0.5 g I2와 1 g KI)을 떨어뜨려 노란 환의 생성 여부로 활성을 확인하였다.
Glucoamylase 활성은 시험관에 2% 가용성 전분 용액 1 mL와 0.2 mL acetic acid를 분주하여 40°C에서 5분간 예열한 후, 효모 배양액 100 μL를 넣고 40°C에서 20분간 반응하였다. 1 N NaOH 용액 100 μL를 첨가해 반응을 정지시켰다. 30분간 실온에 방치한 후, 1 N HCl 용액 0.1 mL를 첨가하여 중화시켰다. 중화된 반응액 1 mL를 glucose assay kit(Sigma-Aldrich Co. Ltd., St. Louis, Mo, USA)를 이용하여 glucose를 정량하였다. 이 반응액을 흡광도(BioTek®, Gen5™, Winooski, VT, USA) 570 nm에서 측정하여 glucoamylase 활성을 산출하였다.
Acidic protease 활성은 시험관에 0.5% casein 용액 150 μL에 Mcllvaine 완충액 100 μL를 넣어 40°C 항온 수조에서 5분간 예열시켰다. 배양액 50 μL를 첨가하여 40°C에서 60분간 반응시킨 후, trichloroacetic acid (TCA) 용액 300 μL를 가해 반응을 정지시켰다. 반응액을 여과하여 침전물을 제거한 뒤, 여과액 100 μL에 0.4 M Na2CO3 용액 500 μL와 페놀시약 100 μL를 넣고 40°C에서 30분간 발색시켰다. 반응 종료액을 흡광도(BioTek®, Gen5™, Winooski, VT, USA) 660 nm에서 측정한 후, acidic protease 활성을 다음 계산식으로 산출하였다.
0.6 mL: Casein 용액 + Mallvaine 용액 + 배양액 + TCA 용액
0.1 mL: 여과액
0.05 mL: 배양액
β-Glucosidase 활성은 Seo 등(2015)의 방법을 변형하여 사용하였다. 10 mM p-nitrophenyl-β-D-glucoside (Sigma-Aldrich Co. Ltd.)를 100 μL 넣고, 100 mM citrate phosphate buffer(pH 4.0)를 890 μL 넣은 후, 배양 상등액을 10 μL 넣고 45°C에서 반응시킨 후, 0.5 M sodium carbonate(Sigma-Aldrich)로 반응을 정지시키고, 원심분리한 상등액을 흡광도(BioTek®, Gen5™, Winooski, VT, USA) 405 nm에서 측정하였다. 분당 생성되는 1 μM은 p-nitrophenol을 1 unit으로 정의하였다. 한편, 정성적인 방법으로 esculin(Sigma-Aldrich Co. Ltd.)을 YPD agar 배지에 0.5%가 되도록 조정하여 멸균한 다음, 1% ferric ammonium citrate(Sigma-Aldrich Co. Ltd.)가 2% 되도록 첨가하여 배지를 제조하였다. 이 배지에 효모를 접종하고 25°C에서 24시간 배양한 후, 주변에 검은 환의 생성 여부로 활성을 평가하였다.
α-Glucosidase 저해능은 탄수화물 식이 후, 혈당 상승을 억제할 수 있어 항당뇨 활성(α-glucosidase inhibition) 측정법으로 분석하였다. 항당뇨 활성은 α-glucosidase activity assay kit (Cat No. MAK123, Sigma-Aldrich Co. Ltd.)에서 negative 활성을 100으로 시료의 활성을 환산하여 저해율을 구하였다. 이러한 활성은 enzyme (0.5 unit/mL α-glucosidase) 50 μL와 효모 배양액 50 μL를 넣고 초기값(A405) initial을 측정하였다. 37°C에서 5분 반응시킨 뒤, 2 mM p-nitrophenyl-α-D-glucopyranoside 50 μL를 넣고, 37°C에서 30분간 반응시킨 후 최종 흡광도(A405 nm) final을 측정하였다. 이때 positive control은 배양 상층액 대신 1% acarbose 50 μL를 넣어 측정하였다. 항당뇨 저해율(%)은 다음과 같은 식으로 계산하였다.
A405 calibrator: 20분 반응 후 calibrator 흡광도 값
A405 water: 20분 반응 후 water 흡광도 값
Angiotensin converting enzyme (ACE) 저해 활성은 ACE inhibition screening kit(Cat No. KT-534, Kamiya Biomedical Co., Seattle, WA, USA)를 사용하여 제조회사의 분석 방법대로 분석하였다. 96 well plate의 blank 1, 2에는 증류수 20 μL와 40 μL씩 분주한 후, 모든 well에 20 μL의 substrate buffer를 처리하였고, blank 2를 제외한 well에는 효소액을 20 μL씩 분주하였다. 37°C 배양기에서 60분간 반응시킨 후, 모든 well에 지시액 200 μL를 처리하여 상온에서 10분간 반응시켜 450 nm의 흡광도에서 측정하였다. 양성 대조군은 ACE 저해제로 널리 사용되는 captopril을 사용하였다. ACE 저해 활성은 아래의 식으로 계산하여 백분율(%)로 나타내었다.
Blank 1: positive control (without ACE inhibition)
Blank 2: reagent blank
Tyrosinase 저해 활성은 tyrosinase inhibitor screening assay kit(Cat No. ab204715, Abcam, Waltham, MA, USA)를 사용하였다. 96 well plate에 시료와 대조군을 각각 20 μL씩 분주하고 각 well에 tyrosinase enzyme 용액 50 μL를 첨가하여 25°C에서 10분 반응시킨 후, tyrosinase 기질을 30 μL를 넣고 510 nm에서 흡광도를 측정하였다. 30-60분간 측정한 타임 포인트 2개를 선택하여 흡광도의 차(ΔA = A2 - A1)에 의하여 다음과 같이 측정했다.
Slope S: 샘플의 흡광도 차
Slope of EC: 효소 대조군(tyrosinase assay buffer)의 흡광도 차
3. 결과 및 고찰
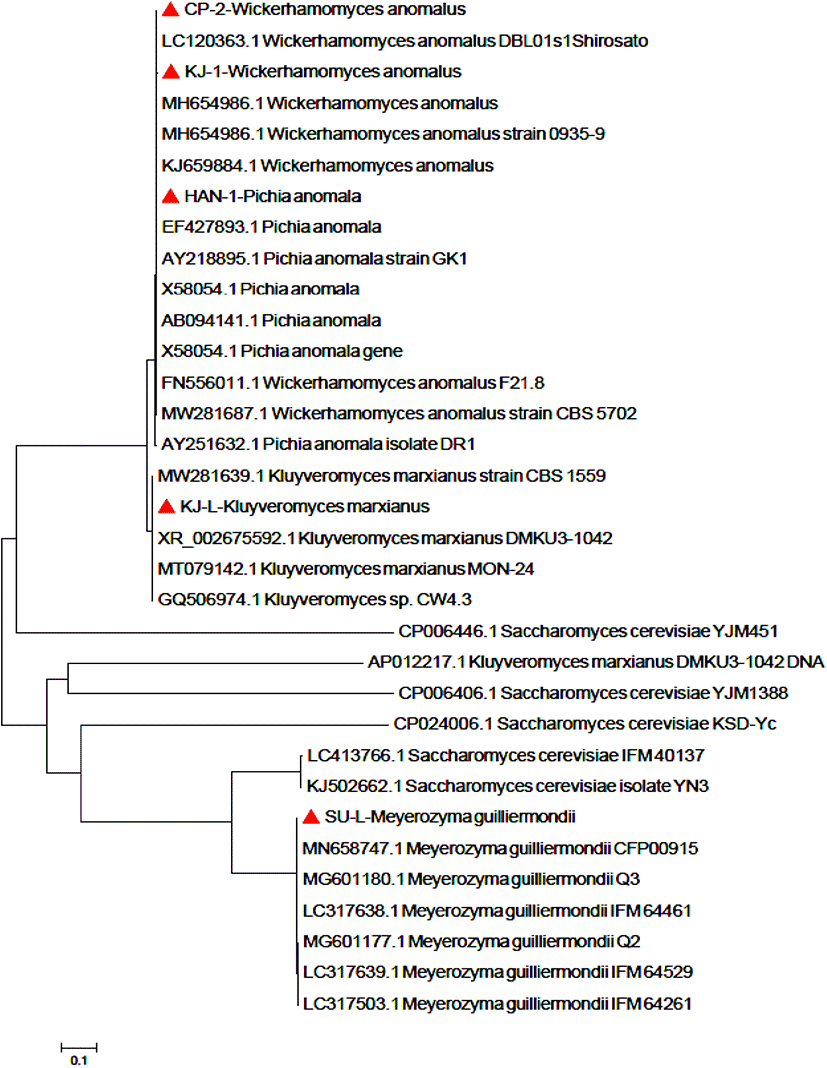
국내 4개 지역(부산, 서울, 전북, 경기)에서 수집한 전통누룩으로부터 순수분리한 5주(KJ-L, KJ-1, CP-2, HAN-1, SU-L)의 효모를 18S rDNA 염기서열 분석을 통해 분자생물학적으로 동정하였다. Saccharomyces cerevisiae (SC)와 non-Saccharomyces cerevisiae (NSC) 효모의 18S rDNA 염기서열을 바탕으로 계통도를 Fig. 1에 나타냈다. 선발된 NSC를 NCBI blast로 염기서열을 분석한 결과, SC 바깥쪽에 Kluyveromyces marxianus, Wickerhamomyces anomalus, Pichia anomalus, Meyerozyma guilliermondii로 각각 동정되었다. NSC로 동정된 5주의 효모는 NCBI에 등록된 type strain과 99% 이상 높은 상동성을 나타내었다.
지역별로 수집한 4종류의 전통누룩에서 분리한 5주 효모의 온도 및 환경적응성(pH 2-pH 9)을 조사한 결과, NSC 4주(K. marxianus KJ-L, W. anomalus KJ-1, W. anomalus CP-2, P. anomalus HAN-1)의 최적 생육 온도는 25°C였다(Table 1). 생육 25°C 대비 35°C에서 18-40%의 생육을 보였고, 저온(5°C와 10°C)에서는 1% 미만에서 7% 이하의 생육을 보였다. 특히, 고온(45°C, 55°C) 조건에서는 이들 효모는 생육하지 않았다. 대조구로 사용한 SC 최적 생육은 25°C 대비 35°C에서 35% 생육과 그 외 온도에서 1% 미만의 생육 특성을 나타낸 것과 비슷하였다(data not shown). 또한, M. guilliermondii SU-L 효모는 35°C가 최적 생육이었으며, 25°C에서는 78%의 생육을 보였다(Table 1). 한편, 분리 효모의 pH에 따른 환경적응성을 조사한 결과, pH 4에서 생육이 가장 양호하였고, 중성인 pH 7보다는 10-60%까지 생육능이 우수하였고, 산성과 알칼리성 조건에서(pH 2, pH 9) 25% 미만으로 NSC 생육능은 저조하였다(Table 2).
선발한 5주의 NSC를 22 °Brix로 보당한 YPD 액체배지에 각각 접종한 후, 25°C에서 5일간 배양하여 알코올 생성량을 조사한 결과, W. anomalus CP-2가 11.73%로 가장 높게 나타났고, K. marxianus KJ-L(11.14%), S. cerevisiae LP(10.54%), P. anomalus HAN-1(10.14%), W. anomalus KJ-1(9.91%), M. guilliermondii SU-L(9.55%) 순으로 알코올을 생산하였다(Table 3). 또한, 이들 효모의 발효에 따른 가용성 당도 변화를 살펴본 결과, 발효가 진행됨에 따라 NSC와 SC는 잔존 당이 영양원이나 발효 기질로 이용되면서 당도는 감소하고 알코올 함량은 증가하였다(Table 3). SC 효모의 고형분 함량은 7.2 °Brix로, NSC 속 효모(7.3–8.3 °Brix)보다 낮게 나타났다. SC의 고형분 함량은 NSC 속 효모보다 낮은 함량을 나타내었는데, 이는 SC가 NSC 속(genus)보다 당을 이용하여 알코올 발효가 더 진행(Baek 등, 2015)된 것과는 상이하게 나타났다. Baek 등(2015)의 보고에서 25°C, 48시간 배양 시, SC의 알코올 생성능은 7.3-9.5%를 보였고, NSC는 2.2-4.7%로 나타낸 것과는 다소 차이를 나타내었다. 그러나 Choi 등(2019)에서는 W. anomalus SC-1이 기존 시판 효모와 비교하여 효과적인 알코올 발효능을 나타낸 것과는 유사하였다. 배양온도와 시간의 차이는 있지만, 대체적으로 SC의 알코올 생성능이 NSC에 비교하여 높다고 알려졌지만, 본 연구에서는 NSC 중에서도 K. marxianus KJ-L과 W. anomalus CP-2가 기존 시판 효모보다 알코올을 5.8-11.3% 더 많이 생산했다. 기존 수입산 양조용 효모 대비 알코올 생성능이 뛰어난 NSC 계열의 효모를 탁·약주 제조에 적극 활용을 권장한다.
일반적으로 NSC는 다양한 효소를 세포 밖으로 분비함으로써 주류의 향기 성분 증가에 큰 영향을 준다고 알려져 있다(Baek 등, 2015). NSC의 알코올 생산에 대한 산업적 활용성을 살펴보고자 세포 밖으로 분비한 다양한 종류의 효소활성을 조사하였다.
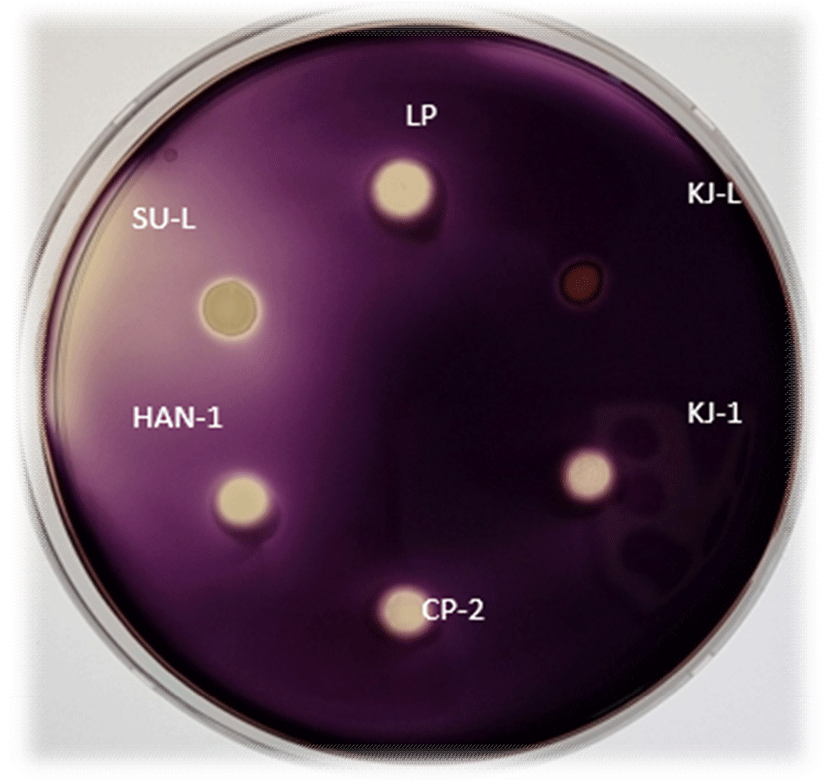
전분은 식물의 광합성 작용으로 생성 및 저장되었다가 amylase에 의해 분해되어 동물의 에너지원으로 사용된다. 분리한 5주 NSC의 요오드 염색법에 따른 전분 분해능을 살펴봤을 때, K. marxianus KJ-L을 제외한 효모는 30°C에서 비슷한 크기의 전분 분해능을 나타냈었다(Fig. 2). 정량적 NSC의 α-amylase 활성 분석에서도 K. marxianus KJ-L 효모의 α-amylase 활성이 가장 낮게 나타났다(Table 4). 본 실험에 대조구로 사용한 SC가 α-amylase 활성이 가장 높았고 W. anomalus CP-2, M. guilliermondii SU-L, P. anomalus HAN-1은 같은 정량값으로 두 번째, W. anomalus KJ-1은 세 번째 순으로 효소활성을 나타냈다. 전통적으로 알코올 발효에 사용되는 효모의 전분 분해력이 낮아 전분 분해에 필요한 α-amylase와 glucoamylase 등의 효소 비용과 그 공정비는 전체 에탄올 생산비에 상당 부분을 차지하고 있다. 특히, α-amylase보다 당화능이 우수한 glucoamylase가 전분분해에 더 필수적이다(Park 등, 1999).
Glucoamylase는 단당류인 glucose 생산에 관여하며, 당화력을 평가하는 주요한 효소(Chung 등, 1990b)로서 배양 상층액을 회수하여 glucose assay kit로 glucose를 정량한 결과, 대조구로 사용한 SC LP(28.24±0.58 units/mL)보다 NSC인 M. guilliermondii SU-L(40.21±1.28 units/mL)이 1.4배 높게 나타났다. 그 외 4종류의 효모는 14.62± 0.14-22.52±2.64 units/mL로 대조구보다 낮은 당화력을 나타내었다(Table 4).
Acidic protease의 경우에는 P. anomala HAN-1이 148.31±23.46 units/mL로 높은 활성을 보였고, M. guilliermondii SU-L은 136.77±16.54로 나타냈지만 대조구인 SC LP(181.77±30.77 units/mL)보다 활성이 낮게 나타내었다(Table 4). 일반적으로 발효식품의 맛과 향기는 주로 미생물이 생성하는 protease에 의해 영향을 받는다고 알려져 있듯이(Lee 등, 1997), 본 실험에서 효모의 protease 활성을 나타내는 정도에 의해 차이가 있으므로 다양한 효모 종균의 자원화로 다양성을 확보할 수 있으리라 기대된다.
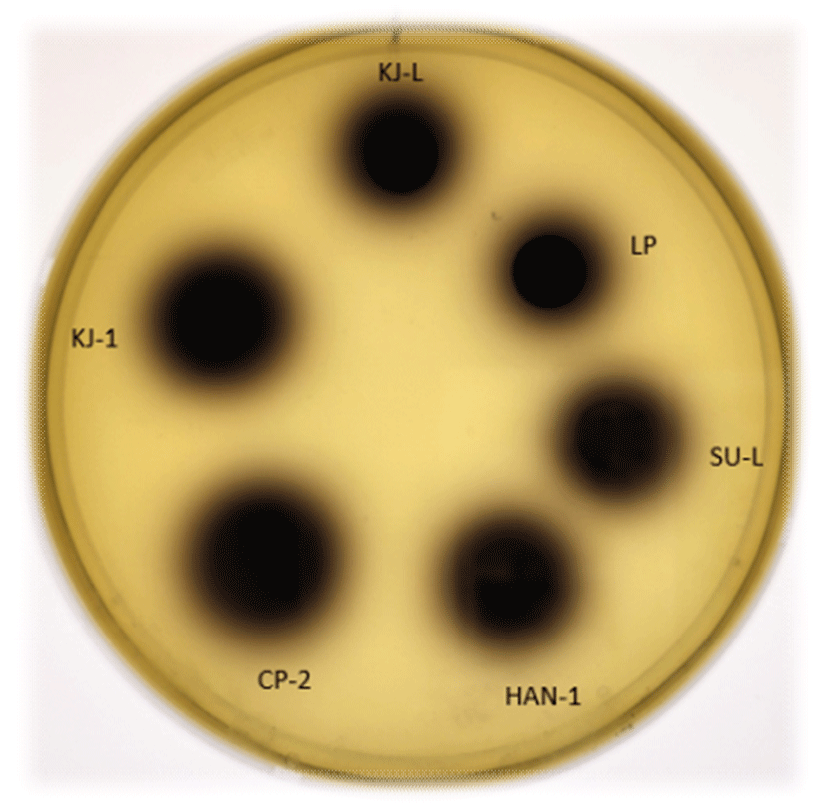
β-Glucosidase 활성은 와인 제조 시, 비휘발성 향기성분을 생산하는 데 있어 중요한 역할을 한다고 알려져 있고(Hernández, 2003), NSC인 Candida stellata, Candida hellenica, Kloeckera apiculata에서 β-glucosidase 활성이 보고되었다(Strauss, 2001). 특히, Hansenispora vinea와 Candida 종이 생산한 glucosidase가 와인의 다양한 향에 영향을 미친다고 알려져 있다(Strauss, 2001). β-Glucosidase 집락 주변의 검은색 환의 생성 여부로 이들 효소의 활성을 구분할 수 있다. 국내 전통누룩에서 분리·선발한 5주의 NSC에서 이들 효소의 활성을 확인하였다(Fig. 3, Table 4). Table 4에 나타낸 것처럼 β-glucosidase 활성은 시판 효모인 SC LP가 3,767.86±128.76 units/mL로 가장 높았으며, P. anomalus HAN-1과 W. anomalus KJ-1, W. anomalus CP-2가 2,910±353.71과 2,386.90± 20.62, 2,172.62±232.37 units/mL 순으로 활성을 나타내었다(Table 4).
이러한 효소활성은 탁·약주 제조에 있어서 Saccharomyces와 non-Saccharomyces cerevisiae를 혼합으로 사용할 경우, 주질의 향미를 증진할 가능성을 시사해 준다.
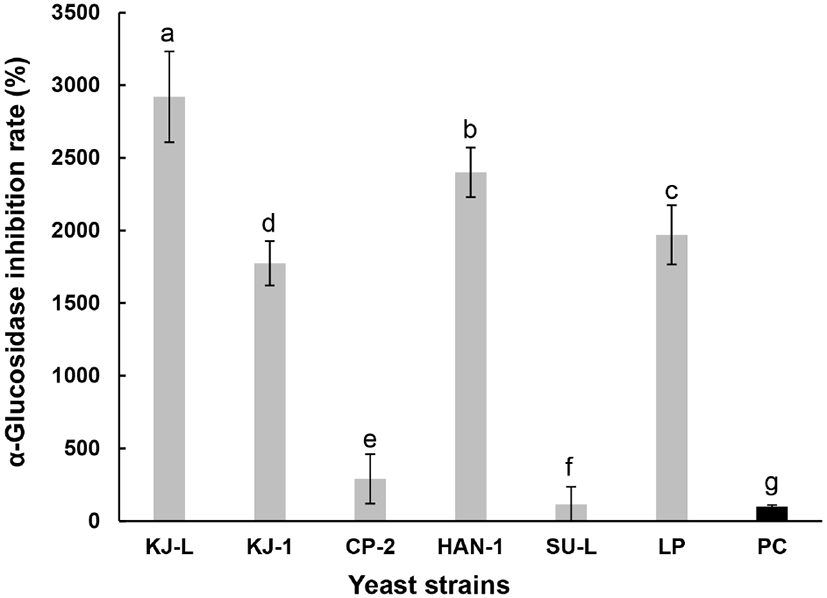
α-Glucosidase는 α-amylase에 의해 분해된 이당류를 최종적으로 단당류로 분해하여 소장의 융모로 흡수되어 혈관으로 이동되고, 효소의 활성 저해는 당질 가수분해와 흡수과정을 지연시킴으로 식후 혈당이 급격히 상승하는 것을 억제할 수 있다(Shinde 등, 2008). Baron(1998)은, α-glucosidase 저해제는 당뇨와 같은 당질 관련 질병을 위한 치료제 개발에 유용하다고 보고하였다. 전통누룩에서 분리한 NSC의 α-glucosidase 저해능을 조사하였고, 그 결과를 Fig. 4에 나타낸 것처럼 K. marxianus KJ-L과 P. anomalus HAN-1 효모가 유의적으로 저해 활성능이 높았다. 경구투여용 α-glucosidase 억제제로 잘 알려진 1% acarbose와의 비교에서도 K. marxianus KJ-L 효모는 약 27배의 높은 활성을 나타냈었다. 이는 Kim(2014)이 보고한 P. burtonii Y257-7의 90.9%보다도 29배 높은 활성을 가지고 있었다.
선발한 5주 효모의 배양액으로부터 얻은 상등액을 취하여 ACE 저해 활성 결과를 Table 5에 나타내었다. ACE는 불활성인 angiotensin Ⅰ을 절단하여 활성형인 angiotensin Ⅱ로 전환하는 촉매 효소로, 혈압을 상승시키는 동시에 혈압 강하 작용을 하는 bradykinin을 분해하는 작용으로 생체 내에서 고혈압을 일으키는 중요한 효소로 알려져 있다(Hong 등, 2004). ACE 저해능을 조사하기 위해 양성 대조군으로 1% captopril(표준물질)로 사용한 결과, 75.81± 0.11%로 나타났으며, NSC의 ACE 저해 활성은 K. marxianus KJ-1, P. anomalus HAN-1, K. marxianus KJ-L, W. anomalus CP-2, M. guilliermondii SU-L 순으로 나타났다. 이는 S. cerevisiae LP보다 1.4-1.7배 높은 활성을 나타낸 것으로 Jang(2012a)이 보고한 P. anomala의 ACE 활성과 같은 결과라 할 수 있다. 우리나라의 서구화된 식습관, 운동 부족, 과도한 스트레스 등 다양한 생활 요인으로 인해 심혈관계 질환 발병률이 높아지고 있는 실정(Kim 등, 2022)에서, 항고혈압 활성을 나타내는 다양한 non-Saccharomyces 계열의 효모를 발효식품에 적용함으로써 합성 고혈압 치료제의 부작용에 대비하여 식품 유래의 ACE 저해 활성물질에 관한 기초 연구로서 향후 산업적 활용성이 기대된다.
Tyrosinase는 인체 내 멜라닌 생합성 경로에서 가장 중요한 초기 속도결정단계에 관여하는 효소로서, 활성이 저해되면 멜라닌 생성을 저해하는 결과를 일으킨다(Kim 등, 2014). 선발한 5주의 NSC 배양 상층액의 tyrosinase 저해 활성 결과를 Table 6에 나타내었다. Kojic acid를 양성 대조군으로 사용하였고, 일반적으로 주류 제조에 가장 많이 사용하는 SC LP와 선발한 NSC의 미백 활성을 조사한 결과, W. anomalus CP-2가 활성(93.18%)이 가장 높았고, W. anomalus KJ-1(86.36%)이 가장 낮았지만, 양성 대조구로 사용한 Kojic acid(63.07%)보다 선발한 효모의 상층액에서 미백 활성이 높은 것을 확인할 수 있었다(Table 6). Jang(2012b)이 보고한 P. anomala의 tyrosinase 저해 활성(3.4%-14.4%)과 비교하여도 선발한 NSC의 tyrosinase 저해 활성이 6-28배 높게 나타났다. 현재, 메디푸드 및 향장산업에서 미백 소재의 이용성이 높아지는 시점에서 본 연구에서 선발한 미백 활성 등이 뛰어난 non-Saccharomyces cerevisiae는 향후, 기능성 발효식품 및 화장품 원료로서 활용성이 기대된다.