
Introduction
Marine algae, also known as seaweed or macroalgae, are photosynthetic eukaryotic organisms that can be found in coastal areas. There are three main seaweed taxa that are classified based on their pigmentation: red algae, green algae, and brown algae (1). Seaweed can survive in harsh environmental conditions because they are able to use numerous strategies to adapt to new challenging environments. To survive in such diverse and extreme environments, seaweed produces a variety of natural bioactive compounds and metabolites, such as polysaccharides, polyunsaturated fatty acids, and phlorotannins (2,3).
Polysaccharides are a complex group of macromolecules that have a wide ranges of bioactivities. These polymers are ubiquitous in nature, occurring in a great number of different organisms, such as mammals and invertebrates. Marine seaweeds are the most important source of non-animal polysaccharides. The molecular structure of these polysaccharides vary depending on the species of seaweed. Based on this, polysaccharides from marine seaweed could be compounds with unique structures and novel biological activities. These polysaccharides have been extensively studied because of their various biological activities including, immunomodulatory, anti-cancer, and antioxidant activities (4-6).
Gloiopeltis furcata (G. furcata) is a red alga belonging to the genus Gloiopeltis of the Endocladiaceae family, that been widely used in traditional herbal medicine, and as a food thickener (7). Polysaccharides with different physiological activities (antitumor, antiviral, anti-inflammatory) have been found in water and ethanol extracts prepared from G. furcata (8,9). However, the physiological activities of G. furcata polysaccharides originating from south Korea, have not been studied in detail.
The biological activity of polysaccharides depends on their structural features such as molecular weight, degree of sulfation, type of sugar, and glycosidic branching. Therefore, when polysaccharides of different molecular weights are involved, changes in biological activity are assured. Moreover, the molecular interactions may be affected by the molecular weights of polysaccharides, there by resulting in different biological properties (10-12).
In this study, the immune-enhancing effect of polysaccharides with different molecular weights from G. furcata were investigated.
Materials and Methods
Gloiopeltis furcata (G. furcata) was purchased from a local supermarket (Daegu, Korea). G. furcata was thoroughly washed with water and air-dried at 50℃. The dried samples were milled using a Wiley-type cutting mill (J-NCM; Jisico Co., Seoul, Korea), sieved (<0.5 mm), and stored at -20℃.
G. furcata (100 g) was suspended in distilled water (10 L), and 3% Viscozyme L (enzyme to substrate ratio) was added. The hydrolytic reaction was performed for 14 h at 60℃. After centrifugation at 3,000 ×g for 30 min (Supra 30K; Hanil Science Industrial Co., Seoul, Korea) to remove debris, the filtrate was concentrated in a rotary evaporator and then precipitated with 5 volumes of 94% ethanol for 24 h at 4℃. The precipitates were acquired after centrifugation and dissolved in distilled water.
Polysaccharides with different molecular weights were identical to the sample used in our previous study (13). Briefly, crude polysaccharide preparation (10 g) was dissolved in 0.1 N HCl (1 L) and maintained at 50℃ for different periods of time. The pH of the solution was then adjusted to 7.0 by the addition of ice-cold 0.1 N NaOH. The Amicon ultra-15 centrifugal filter unit (Merck Millipore, Tullagreen Carrigtwohill, Ireland) used for the separation was made of polysulfone material, which is composed of the pore size with molecular weight cut off (MWCO) 10 kDa and 100 kDa Dissolved crude polysaccharides (15 mL) were placed in a centrifugal filter unit (15 mL capacity) and centrifuged at 3,000 ×g for 30 min (1236MG, Gyrozen Co., Daejeon, Korea). after which they were lyophilized at -70℃ under reduced pressure (Free Zone 2.5, Labconco Co., Kansas, MO, USA).
The RAW264.7 cells (Korean Cell Line Bank, Seoul, Korea) were cultured in DMEM supplemented with 10% fetal bovine serum (WELGENE Inc., Daegu, Korea) and antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin; GIBCO, Rockville, MD, USA) at 37℃ in a humidified atmosphere with 5% CO2 (MCO-18AIC, SANYO Co., Sakata, Japan).
Proliferation of the RAW264.7 cell lines treated with polysaccharides with different molecular weights was evaluated using the MTT (3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide) assay. Briefly, a total of 5×104 cells/well were seeded in to 96-well plates for 24 h, and the cells were treated with various concentrations of the isolated compounds for 24 h. After treatment with each sample, MTT solution, diluted in phosphate buffered saline (PBS, 1 mg/mL final concentration, Sigma-Aldrich Co.), was then added to each well at 1/10 volume of media. The cells were incubated for 4 h at 37℃, and dimethyl sulfoxide was added to dissolve the formazan crystals. The absorbance was measured at 570 nm using a microplate reader (UVM-340, ASYS, Biochrom, Eugendorf, Austria). Proliferation was calculated as follows: Activity (%)=(absorbance sample/ absorbance control)×100.
Nitric oxide (NO) secretion from macrophages and the nitrite concentration were determined by the Griess reaction. RAW264.7 cells were placed in a 96-well plate (5×104 cells/well in a volume of 100 μL) and incubated for 24 h at 37℃. Cultured cells were treated with 100 μL of each sample or LPS (final concentration 0.1 μg/mL), which was used as a positive control. After incubation for 24 h at 37℃, 100 μL of the cultured cell supernatant was mixed with an equal volume of griess reagent (1% sulfanilamide, and 0.1% N-[1-naphthyl] ethylenediamine hydrochloride in distilled water) and left at 25℃ for 10 min. Absorbance at 540 nm was measured using a microplate reader (UVM-340, ASYS Co.). NO secretion from RAW264.7 cells was calculated with reference to a standard curve obtained using NaNO2.
For cytokine determination, RAW264.7 cells were cultured in the presence of each sample or LPS (final concentration 0.1 μg/mL). Cells were incubated for 24 h, and the supernatant was collected and used to determine cytokine production following stimulation by each sample. The production of TNF-α and IL-6 was measured using commercially available ELISA kits (Pepro Tech Inc., Rocky Hill, NJ, USA).
To evaluate the mRNA expression levels of inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2), total RNAs from each samples or LPS (final concentration 0.1 μg/mL) treated RAW264.7 cells were prepared using a total RNA (1 μg) Extraction Kit (Enzynomics Co., Daejeon, Korea). RT-PCR (Mastercycler® Nexus; Eppendorf AG, Hamburg, Germany) was performed using the Revert Aid First Strand cDNA Synthesis Kit (Thermo Scientific, Waltham, MA, USA). Reverse transcriptase amplification was performed by initial denaturation (94℃ for 30 sec), annealing (55℃ for 30 sec), and extension (72℃ for 1 min), followed by a final extension step at 72℃ for 10 min. The products obtained by RT-PCR were separated by electrophoresis (Mupid-ex, Advance, Tokyo, Japan) using a 1.2% agarose gel and stained with Eco View (Daemyung Science Co., Ltd., Seoul, Korea). The gels were then viewed under UV transillumination. The primer sequences (Bioneer, Daejeon, Korea) used in this experiment were as Table 1.
All experiments were performed in triplicate, and means±SD or average values are reported. Duncan’s multiple-range test was used to adjust for multiple comparisons, and null hypotheses were rejected at the 0.05 level. All data were analyzed using SPSS/Windows software (Version 19.0; SPSS Inc., Chicago, IL, USA).
Results and Discussion
In our previous study, we preparation of polysaccharides with different molecular weights from G. furcata, further investigated the chemical compositions (13). The major constituents in molecular weight fractions were total sugar (81.37-85.82%), uronic acid (27.89-32.85 g/100 g), sulfate (33.38-39.04%), and protein (0.35-3.16%), respectively.
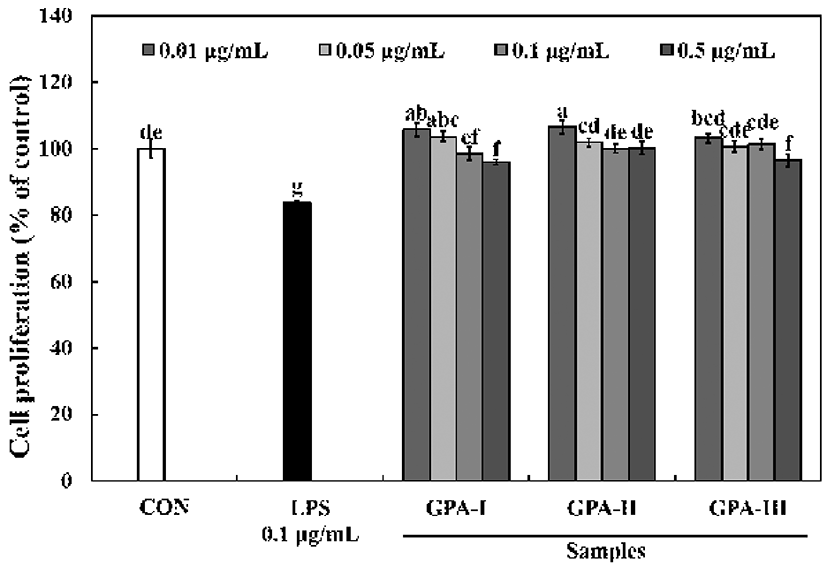
Macrophages play critical roles in host defense, including the phagocytosis of pathogens and apoptotic cells, cytokine production, and the proteolytic processing and presentation of foreign antigens (14). The effect of a crude polysaccharide preparation from G. furcata, and sub-fractions of the same polysaccharide preparation (0.01-0.5 μg/mL) having different molecular weights, on RAW264.7 cell proliferation are shown in Fig. 1. When RAW264.7 cells were cultured with the different sub-fractions of the polysaccharides with different molecular weights, their proliferation increased only slightly, suggesting that these polysaccharides are not cytotoxic to RAW264.7 cells over the range of concentrations tested.
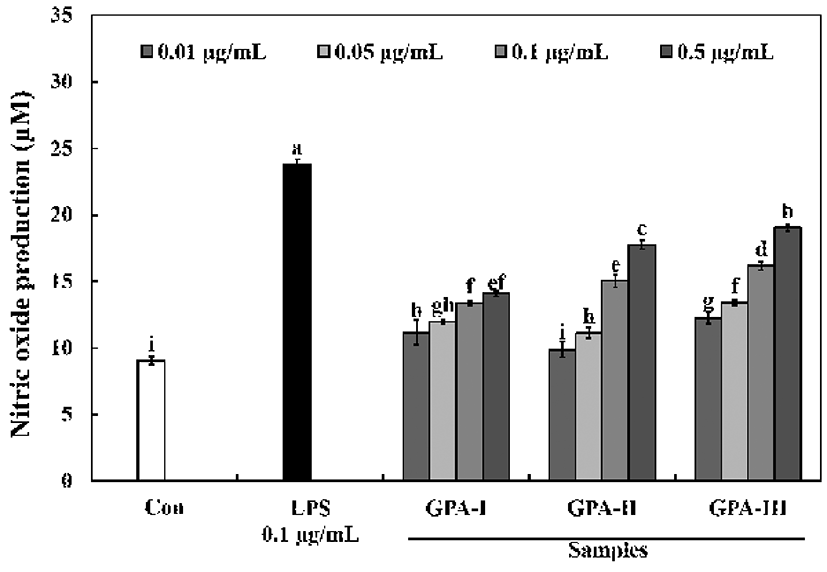
Activated macrophages (RAW264.7) release immunomodulatory factors, such as NO, as well other factors, to remove pathogens and inhibit the growth of invading cells. In particular, the nitric oxide released from macrophages is a highly reactive free radical that plays an important role in the regulation of various immune responses. Therefore, RAW264.7 cells are often used to determine the immunomodulatory activities of various compounds by examining nitric oxide production, as well as their cytokine production profile (15). The polysaccharides with different molecular weights (0.5 μg/mL) were able to significantly stimulate macrophage cells and produce a considerable amount of NO (range of 14.10-19.05 μM), in a dose-dependent manner (Fig. 2). The amount of NO released was higher for polysaccharides of GPA-Ⅲ (≥ 100 kDa) compared to polysaccharides with GPA-Ⅰ (≤10 kDa) or GPA-Ⅱ (10-100 kDa) which resulted in increased levels of NO (range from 12.28-19.05 μM). Generally, NO plays an important role as a vasodilator, neurotransmitter, and in the immunological system as a defense against tumor cells, parasites, and bacteria (16). These results suggest that the sulfate content and the molecular weight of sulfated polysaccharides may contribute to NO production. Moreover, Leiro et al. (17), and Tabarsa (18) have reported that polysaccharides from Ulvarigida and Ulva intestinalis induce high NO production, proportionate to their sulfate contents and MW. These findings suggest that the MW and high sulfate content of polysaccharides are pivotal for its high bioactivity.
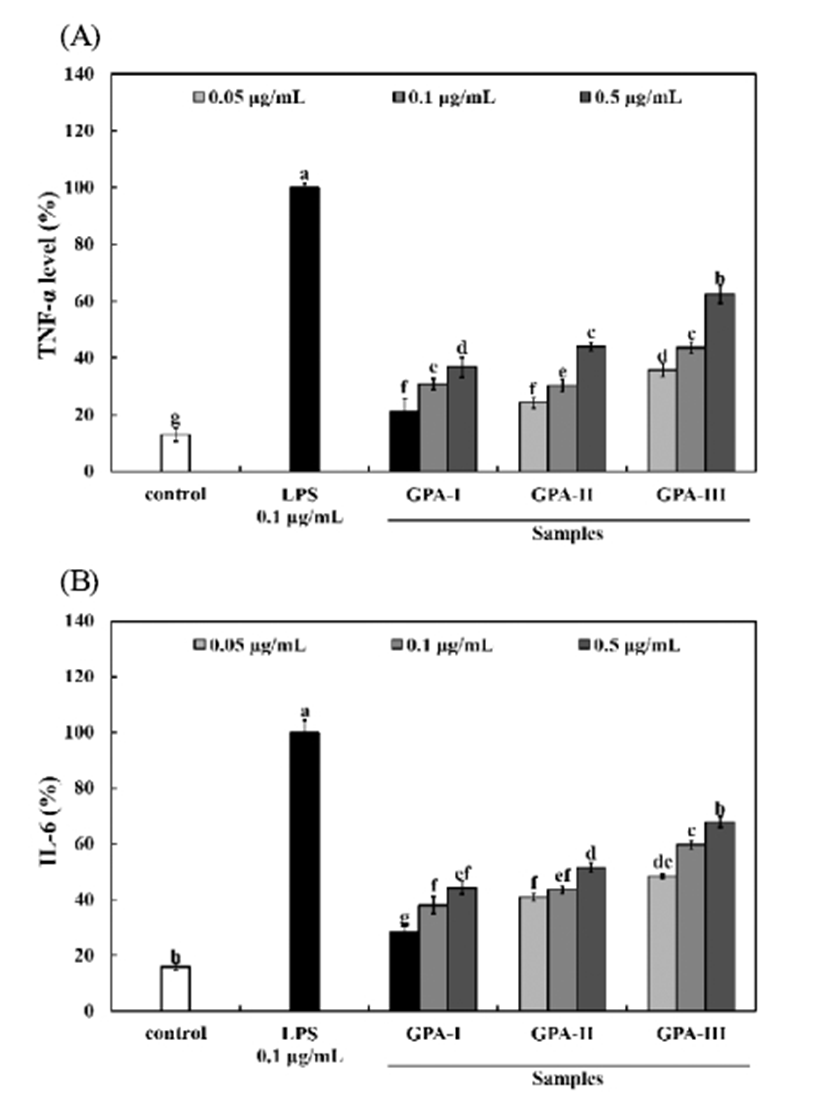
Part of the defense mechanism used by macrophages against pathogens includes the release of pro-inflammatory cytokines such as TNF-α, and IL-6. Polysaccharides with a GPA-Ⅲ (≥100 kDa) stimulated the production of TNF-α (35.77-62.37% of positive control), and IL-6 (48.42-67.87% of positive control) from RAW264.7 macrophages to levels that were 2-4 times greater than the negative control (13.02%, and 15.85% of positive control). These increases in both TNF- α, and IL-6 secretion from RAW264.7 cells occurred in a dose-dependent manner (Fig. 3). These data suggest that polysaccharides with different molecular weights have a beneficial effect on immune stimulation. Cytokines are signaling molecules that control homeostasis of the organism by regulating cell differentiation, proliferation, and apoptosis, as well as defense functions such as immune responses and inflammatory reactions. Cytokines are produced by cells that possess regulatory properties and are found very close to, or in direct contact with, target cells. By binding to specific cell surface receptors, they can exert their effects on cell proliferation, differentiation, as well as exerting other effects. Once released into the blood they behave like hormones and affect different tissues and organs, often distant from the original site of release (19). TNF-α is a cytokine that has both antitumor and immunoregulatory functions. It has been recognized to be an important host defense cytokine that can affects tumor cells. IL-6 is also considered to be an important mediator of acute inflammatory responses and functions as a differentiation and growth factor for hematopoietic precursor cells, B cells, T cells, keratinocytes, neuronal cells, osteoclasts, and endothelial cells (20).
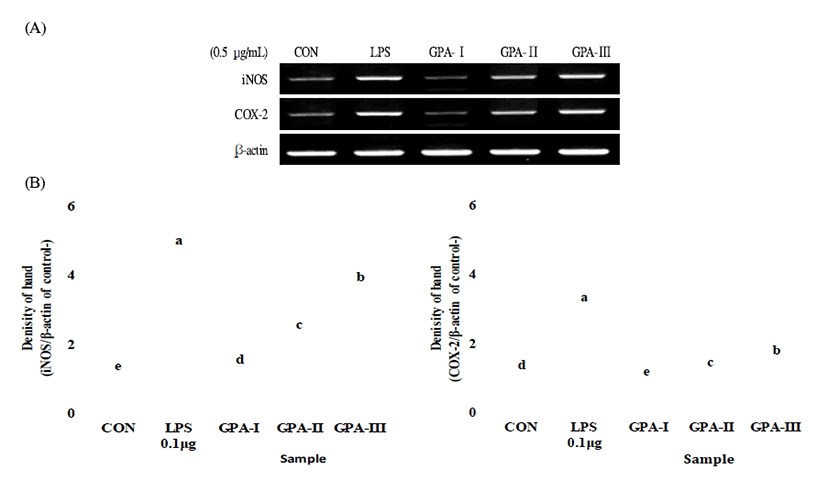
Although host defense mechanisms are complex, it is known that macrophages are key participants in the innate immune response to the invasion of pathogenic organisms (21). RAW264.7 macrophage cells were treated with the different subfractions of polysaccharides with different molecular weights, and RNA was harvested from the cells and subjected to RT-PCR. As shown in Fig. 4, the sulfated polysaccharides with different molecular weights stimulated the induction of the mRNAs encoding iNOS and COX-2. Moreover, the data revealed that polysaccharides with a molecular weight ≥100 kDa might be immunostimulatory polysaccharides that could activate macrophages. The sulfated polysaccharides with a GPA-Ⅲ (≥100 kDa) also stimulated the production of the effector molecule NO in macrophages. It has been found that NO inhibits the early stages in viral replication, and thus prevents viral spread, promoting viral clearance and recovery of the host (22). These results were supported by studies by Lee et al. (23), who demonstrated that sulfated galactans from Codium fragile had macrophage stimulating effects.
요 약
본 연구에서는 불등풀가사리(홍조류) 다당류를 기능성 소재로의 활용 가능성을 확인하기 위하여 분자량이 다른 다당류 3종을 제조하여 다양한 면역증강 효과를 확인하였 다. GPA-Ⅲ(≥100 kDa)는 nitric oxide 및 cytokine(TNF-a 및 IL-6)분석에서 농도 유의적으로 함량이 증가하였으며, RT-PCR 분석을 통한 유전자 발현에서 iNOS 및 COX-2에서 높은 발현을 유도하여 면역증강 효과를 확인하였다. 따라 서, GPA-Ⅲ는 기능성식품 및 의약품의 잠재적 응용 가능성 을 가지는 면역 증강제임을 나타내었다.