
1. 서론
발효(fermentation)는 효모나 세균 등의 미생물이 유기화합물을 분해하여 이산화탄소 및 알코올류, 유기산류를 생산하는 과정이다(Lee, 2019). 발효 과정으로 만들어진 식품은 다양한 성분이 합성되므로 독특한 향과 맛을 지니게 되며, 특히 한국의 전통음식인 김치나 청국장은 대표적인 발효식품 중 하나로 장 건강에 매우 유익하게 작용한다(Kim 등, 2018). 최근 세균, 곰팡이, 효모 등 다양한 미생물을 활용한 여러 가지 발효기술은 항산화, 항균 및 항암 등의 생리활성 효과를 증대시키는 것으로 밝혀졌으며, 특히 유산균을 활용한 천연물 발효기술은 천연물 유래 생리활성 성분 및 기능성의 증가를 쉽게 극대화할 수 있다는 이점이 있어 다양하게 이용되고 있다(Lee 등, 2019).
Lactobacillus plantarum(L. plantarum)은 유산균이자 젖산균에 속하는 세균으로 그람 양성의 간균이며, 많은 발효식품 및 혐기성 식물 물질에서 발견된다(Zheng 등, 2020). Choi 등(2021)은 국내산 꾸지뽕 나무 열매 추출물과 L. plantarum을 이용한 꾸지뽕 나무 열매 발효 추출물의 생리활성을 비교한 결과, DPPH 및 ABTS radical 소거 활성 등 항산화 활성이 발효하지 않은 열매 추출물보다 약 4배 정도 증가하였음을 밝혔고, 폴리페놀 및 플라보노이드 함량이 증가하는 것을 확인하였다. 또한, Song 등(2009)은 김치에서 분리한 L. plantarum이 식중독 원인균에 대한 항균 활성을 갖는다고 하였으며, Choi 등(2020)은 해당 균주로 발효한 뽕잎 추출물이 우수한 항당뇨 효과를 지닌다고 보고하였다. Lactobacillus brevis(L. brevis)는 그람 양성의 젖산균으로 장 건강에 유익하게 작용하는 프로바이오틱스로 알려져 있다. Park(2014)은 L. brevis 균주를 이용한 오미자 당침액의 발효 특성을 연구한 결과, 발효물의 우수한 항산화 활성과 항고혈압 활성을 보고하였으며, Song과 Park(2018)은 해당 균주를 이용한 발효 천마의 경우 총폴리페놀 함량의 증가는 물론 라디칼 소거능이 증가하였으며, 식중독균에 대한 항균 활성이 높아졌음을 보고하였다.
작두콩(Canavalia ensiformis)은 콩과 한해살이 덩굴성 한해살이풀로 열대아시아, 아프리카, 인도 열대지방이 원산지이며, 모양이 작두를 닮았다고 하여 작두 또는 도두(刀豆)라고 불린다(Joo 등, 2001). 종실뿐만 아니라 뿌리, 꼬투리, 잎 등 식물체 모두 식용 및 약용으로 사용하며, 축농증, 치질, 종기 등 화농성 염증 등에 한약 소재로 사용하는 등 민간요법에 주로 사용되었다. 현재 국내에서는 일부 지역에서 생산되고 있으며, 정확한 생산량에 대한 통계는 존재하지 않으나 여러 가지 생리활성이 확인됨에 따라 관심이 증대되면서 생산량이 증가하는 추세이다.
작두콩의 생리활성은 작두콩에 함유된 약용성분인 canavanine, canavalia, hemaglutinine 등에 의한 것으로 확인되었으며, 이러한 약용성분에 따라 항산화, 항균, 항암, 항염증, 혈액순환 등의 다양한 건강 기능성이 입증되었다(Chung 등, 2014; Faheina-Martins 등, 2012; Kim 등, 2013a). 그러나 작두콩 발효에 의한 생리 기능성에 관한 연구는 극히 미미하며, 청국장 발효균인 Bacillus 속 미생물에 의한 생리 기능성에 제한되어 있다(Choi 등, 2021). 이에 따라 유산균인 Lactobacillus 속을 이용한 작두콩 발효액을 제조하고, 이의 항산화, 항염증, 항균 활성 등의 생리기능성을 조사하여 기능성 식품 소재 개발을 위한 기초자료로 활용하고자 한다.
2. 재료 및 방법
본 실험에 사용된 작두콩은 2022년 5월에 (주)황금농원식품에서 친환경(무농약)으로 재배하여 분말화한 것을 구입하였으며, 4°C 냉장고 온도에서 보관하면서 실험에 사용하였다. 발효액 제조를 위해 작두콩 분말과 흑설탕을 1:1(w/w) 비율로 혼합한 혼합물에 1:4(w/v)의 비율로 멸균증류수를 첨가하고 고압멸균기(HR-S-2000, Ilshin Autoclave Co., Daejeon, Korea)를 이용하여 멸균하였다. L. plantarum (KCTC 13093) 및 L. brevis(KCTC 13094)는 생물자원센터(Korean Culture Center of Microorganisms, Daejeon, Korea)에서 구입하였으며, 37°C MRS 배지에서 24시간마다 총 3회 계대배양한 것을 활성화 균주로 사용하였다. 이를 작두콩 분말과 흑설탕 혼합물에 5%(v/v) 비율로 접종하여 3주간 발효하였으며, 발효 후 원심분리를 통해 상등액을 0.45 μm의 membrane filter (Millipore Co., Bedford, MA, USA)로 여과하여 생리활성 측정에 이용하였다.
총폴리페놀 함량은 Folin-Ciocalteau’s 방법(Singleton과 Rossi, 1965)을 약간 변형하여 측정하였다. 즉 시료 50 μL에 2 N Folin-Ciocalteau’s 시약을 25 μL 첨가한 후, 20% Na2CO3 150 μL를 추가로 첨가하고 실온에서 15분간 방치하였다. 이후 ELISA plate reader(VersaMax, Molecular Devices Corporation, Sunnyvale, CA, USA)를 이용하여 725 nm에서 흡광도를 측정하였다. Tannic acid(Sigma-Aldrich Chemical Co., St. Louis, MO, USA)를 사용하여 표준 검량선을 작성하였으며, 이를 이용하여 작두콩 발효액의 총폴리페놀 함량을 μg/mL로 계산하여 나타내었다. 총플라보노이드 함량은 Davis법(1947)을 변형하여 측정하였다. 발효액 20 μL와 diethylene glycol 200 μL를 혼합한 후 1 N NaOH를 20 μL 첨가하여 40°C의 항온수조에서 1시간 동안 반응시켰다. 이것을 420 nm에서 흡광도를 측정하였으며, quercetin(Sigma-Aldrich Chemical Co.)를 사용하여 표준 검량선을 작성하였다. 작두콩 발효액의 총플라보노이드 함량은 μg/mL로 계산하여 나타내었다.
작두콩 발효액의 항산화 활성은 DPPH 라디칼 소거 활성으로 Blois법(1958)에 따라 측정하였다. 멸균증류수를 이용하여 100%, 50%, 25%, 12.5%의 농도로 제조한 발효액을 시료로 하였으며, 시료 100 μL에 200 μM의 1,1-diphenyl-2-picrylhydrazyl(DPPH) 용액(Sigma-Aldrich Chemical Co.) 400 μL를 넣고 혼합한 다음 실온에서 10분간 반응시킨 후 ELISA plate reader(VersaMax, Molecular Devices Co.)를 이용하여 525 nm에서 흡광도를 측정하였다. DPPH 라디칼 소거 활성은 대조군과 실험군과의 흡광도 차이를 비교하여 나타내었다.
작두콩 발효액의 항염증 활성은 nitric oxide(NO) 소거 활성으로 측정하였다. 대식세포인 RAW 264.7 세포는 한국세포주은행(Korea Cell Line Bank, Seoul, Korea)에서 분양받아 사용하였으며, dulbeco modified eagle’s medium(DMEM, Welgene, Gyeongsan, Korea)에 10% fetal bovine serum(FBS, Welgene)과 1% penicillin-streptomycin(Welgene)을 첨가하여 37°C, 5% CO2 incubator에서 배양하였다. 배양한 RAW 264.7 세포를 1×104 cell/well 밀도로 96 well plate에 분주한 후, 24시간 동안 안정화시키고 starvation을 위해 serum-free medium으로 바꿔주었다. 이것을 4시간 동안 배양한 후, 1 μg/mL 농도의 lipopolysaccharides(LPS)(Sigma-Aldrich Chemical Co.)와 시료를 첨가하고 18시간 배양하였으며, 상등액을 griess reagent(Sigma-Aldrich Chemical Co.)와 1:1(v/v) 비율로 섞어서 15분간 반응시킨 후 540 nm에서 흡광도를 측정하였다. NO 소거 활성은 대조군과 실험군과의 흡광도 차이를 비교하여 나타내었다.
항염증 인자인 iNOS와 COX2에 대한 발현 정도를 확인하기 위해 RT-qPCR을 진행하였으며, 방법은 다음과 같다. RNA는 1 μg/mL의 LPS와 발효액을 처리한 RAW 264.7 세포로부터 GeneJET RNA purification Kit(Thermo Fisher Scientific, Waltham, MA, USA)를 이용하여 추출하였다. 이를 reverse transcription system(Promega Co., Medison, WI, USA)을 이용하여 cDNA를 합성하고 이것을 사용하여 qPCR을 진행하였다. 이때 사용된 qPCR은 Tapman Universal PCR Master Mix(Applied Biosystems, Foster City, CA, USA)이었으며, GAPDH (Mm99999915_g1), iNOS(Mm00440502_m1), COX2 (Mm03294838_g1) probe가 사용되었다.
작두콩 발효액의 항균 활성은 디스크확산법(paper disk)을 이용하여 측정하였다. 항균 활성을 측정하기 위해 사용한 균주인 녹농균(Pseudomonas aeruginosa KCTC 22074), 포도상구균(Staphylococcus intermedius KCTC 3344, Staphylococcus haemolyticus KCTC 3341, Staphylococcus epidermidis KCTC 3958), 대장균(Escherichia coli KCTC 2571), 효모(Candida albicans KCTC 7729)는 생물자원센터(Korean Culture Center of Microorganisms)에서 구입하였으며, 24시간 배양한 후 각 균에 맞는 배지에 도말하고 준비된 paper disk에 대조군 및 발효액을 완전히 흡수시켰다. 균주의 배양조건에 따라 24시간 배양하였으며, 미생물이 자라지 않는 부분(clear zone, mm)의 diameter를 측정하여 항균력을 나타내었다.
작두콩 발효액의 세포 독성은 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide(MTT)법을 약간 변형하여 측정하였다. RAW 264.7 세포(Korea Cell Line Bank)를 1×104 cell/well 밀도로 96 well plate에 분주한 후, 24시간 동안 안정화시키고 시료 2.5, 5, 10 μL를 첨가한 후 48시간 배양하였다. 배양 후, CellTiter 96®AQueous One Solution Cell Proliferation Assay kit(Promega Co.)를 사용하여 측정하였다.
작두콩 발효액의 총폴리페놀 및 플라보노이드 함량과 항산화 및 항염증, 항균, 세포 독성 실험은 3회 이상의 반복으로 수행되었으며, 결과는 평균과 표준편차(mean±standard deviation)로 나타내었다. 이것을 토대로 통계분석을 실시하였으며, SPSS(Ver. 26.0, SPSS Inc, chicago, IL, USA) 통계 프로그램을 사용하여 일원배치 분산분석(one-way ANOVA)을 실시한 후 Duncan’s multiple range test로 유의성(p<0.001, p<0.01, p<0.05)을 검증하였다.
3. 결과 및 고찰
폴리페놀 및 플라보노이드는 식용 가능한 콩에서의 주요 천연 항산화 물질이다. 폴리페놀류 화합물에 포함된 다량의 하이드록실기(-OH)가 다른 물질들과 수소를 공유하며 결합하기 때문에 자유 라디칼과 같은 활성산소종을 효과적으로 제거해준다(Herrmann과 Nagel, 1989; Yusof 등, 1990). 이에 따라 발효 후 작두콩의 총폴리페놀 및 플라보노이드 함량을 측정하였으며 결과는 Table 1과 같다. 작두콩 발효액의 총폴리페놀 함량은 비 발효 작두콩 분말의 총폴리페놀 함량에 비해 유의적으로 낮았다(p<0.05). Lactobacillus 속 균주의 종에 상관없이 모두 발효 후 낮은 함량을 보였으며, 이는 발효로 다양한 폴리페놀류가 분해되어 나타난 결과로 생각된다. 총플라보노이드 함량 또한 작두콩 발효액이 비 발효 작두콩 분말보다 유의적으로 낮은 함량을 보였다(p<0.05). 총폴리페놀 함량과 같은 경향을 보였으며, Lactobacillus 속 균주의 종에 상관없이 모두 발효 후 낮은 함량이 측정된 것으로 보아 발효에 따른 항산화 효과는 없을 것으로 예상된다. Park 등(2018c)은 아마씨를 유산균으로 발효하여 추출한 추출물의 총폴리페놀 함량을 측정한 결과, 발효 아마씨 추출물은 전반적으로 발효하지 않은 대조군에 비해 약 1.5-8배 증가된 결과를 보였다고 보고하였다. 반면 Adebo 등(2018)은 Lactobacillus fermentum에 의해 발효된 수수의 총폴리페놀, 총플라보노이드 함량을 측정한 결과, 오히려 발효 전보다 감소하였으며, 이는 생리활성 화합물의 가수분해에 의한 것이라고 보고하였다. Yang 등(2022)은 찔레나무 열매 젖산 발효물의 총폴리페놀 함량을 측정한 결과 발효 시간에 따라 증감에 차이를 보였으며, Ahn 등(2021)은 아티초크 유산균 발효물의 발효 전후 총폴리페놀을 측정한 결과, 발효 균주에 따라 증가하거나 감소하는 경우가 있었다고 보고하였다. 즉 발효에 따른 작두콩의 총폴리페놀 및 플라보노이드 함량이 감소한 것은 생리활성 화합물이 가수분해되어 나타난 결과로 생각되며, 이는 발효 시간, 균주 등 발효 조건에 따라 차이가 있을 것으로 생각된다.
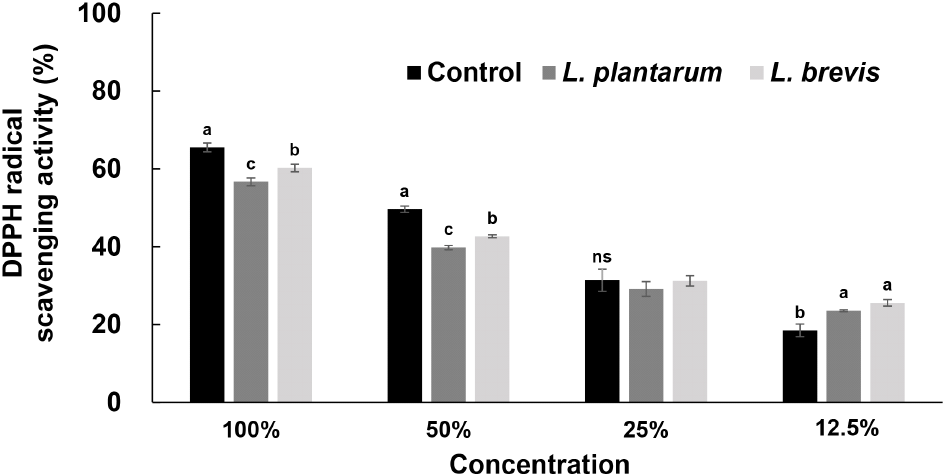
DPPH는 질소를 중심으로 구성된 안정한 라디칼 화합물로 피부노화, 면역기능 억제, 염증 유발, 기미, 주근깨 등의 원인이 되는 활성산소이다(Ammar 등, 2009). 이러한 활성산소는 항산화 물질에 의해 제거되거나 감소하므로 항산화 활성을 측정하기 위한 in vitro 방법으로 라디칼 소거 활성이 주로 사용되며, 그중에서도 특히 DPPH 라디칼 소거 활성은 비교적 간단하고 비용이 저렴하여 항산화 활성을 평가하는데 대표적으로 사용된다. 이에 따라 DPPH 라디칼 소거 활성으로 항산화 활성을 평가하였으며, 그 결과는 Fig. 1과 같다. 발효액을 농도별로 처리하여 DPPH 라디칼 소거 활성을 측정한 결과 농도 의존적인 경향을 보였으며, 작두콩 발효액은 12.5% 발효액을 제외하고 모든 실험군에서 비 발효 작두콩보다 낮은 활성을 보였다. 25% 발효액의 경우 비 발효 작두콩과 유의적인 차이는 없었으며, L. plantarum에 의한 발효액보다 L. brevis에 의한 발효액의 항산화 효과가 조금 더 높았다. Kim 등(2013b)은 발효 미생물에 따른 인삼꽃의 항산화 활성을 측정한 결과 L. plantarum으로 발효한 인삼꽃의 DPPH 라디칼 소거 활성의 경우 무발효 추출물인 control보다 약 2배 정도 낮은 활성을 보였다고 보고하여 본 연구 결과와 유사하였으며, 이러한 경향은 폴리페놀 함량과 깊은 상관관계를 보였다고 하여 본 연구 결과와 일치하였다. Kang 등(2020) 또한 미생물 발효에 의한 구멍갈파래의 항산화 활성을 측정한 결과, Candida utilis 균주로 발효시킨 발효물의 DPPH 라디칼 소거능은 무발효물보다 낮았다고 보고하여 본 연구 결과와 유사하였다. 이는 총페놀 함량과 상관관계가 있으며, 균주에 따른 차이가 있는 것으로 보고되어 본 연구 결과 또한 총폴리페놀 및 플라보노이드 함량과 깊은 상관관계가 있고, 어떤 균주로 발효하느냐에 따른 차이도 있을 것으로 생각된다. 즉, 작두콩은 발효에 의한 항산화 활성 효과는 없는 것으로 판단되나, 낮은 농도에서는 발효로 인한 항산화 효과가 나타나는 것으로 보아 이에 대한 추후 연구가 필요할 것으로 생각된다.
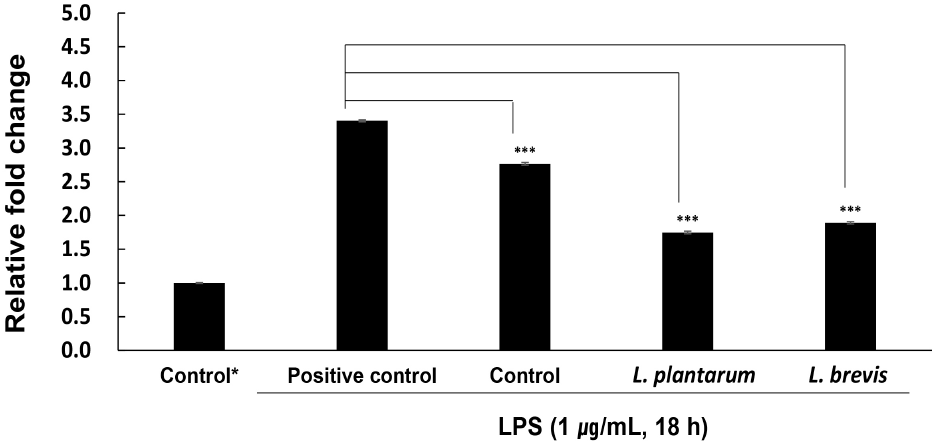
체내에 생성된 NO가 정상적인 범위 내에서는 면역 조절, 혈관 확장, 신경 전달 등 신진대사 작용에 도움이 되지만 염증반응 과정에서 생긴 다량의 NO는 염증매개체를 활성화함에 따라 세포 및 조직에 있어 산화적 손상과 염증 등의 질병을 유발한다(Wyszyńska 등, 2022). 이에 따라 세포에 LPS로 염증을 유발한 후 작두콩 발효액을 처리하여 억제된 NO량으로 항염증 활성을 평가하였으며, 그 결과는 Fig. 2와 같다. LPS를 처리한 결과 NO의 생성이 control에 비해 약 3.4배 증가하여 염증이 유도된 것을 확인하였으며, 시료를 처리한 실험군에서 NO의 생성이 유의적으로 낮아지는 결과를 보였다(p<0.001). 균주를 접종하지 않은 대조군과 L. plantarum 및 L. brevis 균주로 발효한 발효액의 relative fold 값은 각각 2.764와 1.747 및 1.891로 L. plantarum로 발효한 발효액의 NO 억제률이 가장 높았으며, 균주 간 유의적인 차이는 없었다(p>0.05). 즉, 작두콩은 발효 후 NO의 생성이 급감하였으며, 균주의 종류에 유의적인 차이가 없는 것으로 보아 두 균주의 차이 없이 발효로 항염증 효과가 크게 증가하였음을 알 수 있었다. Kwon 등(2015)은 발효톳 추출물의 항염증 활성을 측정한 결과, 균주를 접종하지 않은 무발효물보다 Weissella sp. SH-1과 L. casei 접종군에서 더 높은 활성을 보였으며, Weissella sp. SH-1보다 L. casei를 접종한 발효추출물에서 NO 생성 억제능이 더 우수하였음을 확인하여 균주에 따른 차이가 있음을 증명하였다. Lee 등(2022) 또한 양배추 유산균 발효로 인해 염증을 매개하는 NO의 생성량이 감소하였음을 확인하였으며, Park 등(2018a)의 연구에서도 유산균인 L. plantarum에 의해 층꽃나무 발효물의 항염증 능력이 향상되었다고 보고하여 본 연구 결과와 일치하였다.
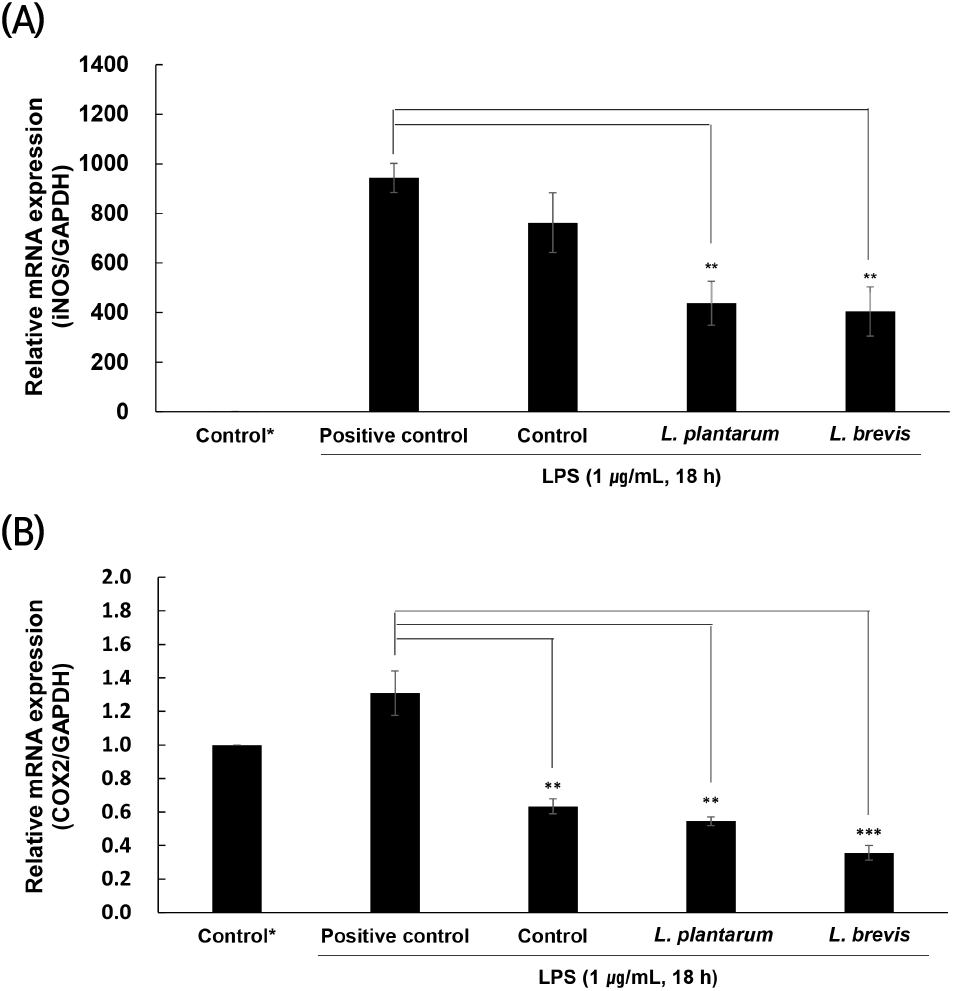
NO의 생성은 각각 iNOS와 COX2에 의해 생성되므로 염증 인자인 NO의 생성을 조절하는 iNOS와 COX2의 조절은 항염증 관련 타켓으로 중요하다고 여겨지고 있다(Park 등, 2018b). 이에 따라 작두콩 발효액의 iNOS와 COX2의 유전자 발현 정도를 측정하였으며, 결과는 Fig. 3에 나타내었다. iNOS의 경우(Fig. 3(A)), LPS 처리 후 급격히 증가하여 염증이 유발된 것을 확인하였으며, 균을 접종하지 않은 대조군은 positive control과 유의적인 차이가 없었으나, 균을 접종한 발효액의 경우에는 유의적으로 iNOS가 감소한 것을 확인하였다(p<0.01). L. plantarum에 의해 발효된 발효액의 iNOS 발현량은 437.40±89.20, L. brevis에 의해 발효된 발효액의 iNOS 발현량은 403.81±99.51로 L. brevis에 의해 발효된 발효액이 L. plantarum에 의해 발효된 발효액보다 높은 활성을 나타내었다. COX2의 경우(Fig. 3(B)), 균을 접종하지 않은 대조군과 균을 접종한 발효액 모두 positive control보다 유의적으로 낮은 발현량을 보였으며(p<0.05), 특히 L. brevis에 의해 발효된 발효액이 가장 낮은 COX2의 발현량을 가짐으로써, 활성이 가장 높은 것을 알 수 있었다. 이러한 결과, 작두콩 발효액은 iNOS와 COX2 발현 조절을 통해 NO의 생성을 억제하는 것으로 판단되며, 이는 항염증 소재로써 충분한 가능성을 보여주는 것으로 생각된다.
작두콩 발효액의 균주에 대한 항균 활성 결과는 Table 2와 같다. 비 발효 작두콩을 대조군으로 하여 disc 주위의 생육저해환(clear zone)의 크기(mm)를 비교한 결과, 비 발효 작두콩에 비해 발효 작두콩의 생육저해환 크기가 유의하게 높은 것으로 확인되었으며, 이러한 결과로 발효 후 항균 활성이 증가하였음을 알 수 있었다. 그러나 효모인 Candida albicans(C. albicans)에 대한 항균 활성이 없는 것으로 보아 균주에 따른 차이를 보였고, 녹농균, 포도상구균, 대장균 또한 각 균주에 따른 생육저해환의 크기가 다른 것으로 보아 활성에 차이가 있는 것으로 보인다. 특히 작두콩 발효액은 Pseudomonas aeruginosa(P. aeruginosa) 즉, 녹농균에 대한 저해 활성이 L. plantarum으로 발효한 발효액이 30.0±0.0 mm, L. brevis로 발효한 발효액이 26.0±0.0 mm으로 가장 우수하였다. P. aeruginosa는 그람 음성의 호기성 간균에 속하는 균으로, 균 자체로는 병원성이 약하지만 면역이 저하된 개체에서는 다른 균종과 함께 작용하여 기회 감염의 주요 원인이 된다(Seo, 2007). 일반적으로 각막염이 이러한 녹농균에 의해 유발되며, 최근 미세먼지와 렌즈 착용으로 인한 결막염, 각막염 등의 발생율이 증가 추세에 있기 때문에 더욱 중요한 세균이다. Lee 등(2020)은 목련 에센셜 오일의 항균 활성을 측정한 결과, P. aeruginosa에 대한 항균 효과로 10 mg의 농도에서 14 mm의 clear zone을 확인하였다고 보고하였다. 또한, Choi 등(2019)은 청귤의 항균 활성을 측정한 결과, P. aeruginosa에 대하여 5 mg과 10 mg의 청귤 주스를 처리한 결과, clear zone의 크기가 각각 11 mm, 15 mm로 확인되었다고 보고하여 작두콩 발효액의 항균 효과가 더 높았음이 증명되었다. 작두콩의 Staphylococcus sp. 즉, 포도상구균에 대한 억제능은 녹농균에 비해 낮은 저해능을 보였으나, L. plantarum에 의한 발효액이 L. brevis에 의한 발효액보다 높은 활성을 나타낸 경향은 일치하였다. 대장균, Escherichia coli O157:H7에 대한 억제능은 녹농균과 포도상구균에 비해 낮은 활성을 보였으며, C. albicans에 대한 효과는 없는 것으로 보아 균주에 대한 차이가 있는 것으로 보인다. 포도상구균은 그람 양성의 구균으로 대표적인 식중독균이며, 사람에게 패혈증 등의 질병을 일으킨다(Newell 등, 2010). C. albicans는 칸디다증의 원인이 되는 병원성 곰팡이로 피부 점막 감염부터 모든 기관을 침범할 수 있는 병원체로 잘 알려져 있다(Pappas 등, 2004). 이로써 작두콩 발효액의 항균 효과는 균주에 따른 차이는 있으나 발효로 인한 저해능이 증가하였음을 확인하였고, 해당 균주에 의한 각종 감염에 의한 질병을 예방할 수 있을 것으로 생각된다.
4. 요약
본 연구는 발효에 의한 생리기능성을 조사하여 기능성 식품 소재 개발을 위한 기초자료를 제공하고자 Lactobacillus 속을 이용한 작두콩 발효액을 제조하여 항산화, 항염증, 항균 활성을 평가하였다. 총폴리페놀 및 플라보노이드 함량은 균주의 종류에 상관없이 발효 후 유의적으로 낮은 결과를 보였으며(p<0.05), 항산화 활성을 측정한 DPPH 라디칼 소거 활성 또한 발효에 따른 효과가 나타나지 않았다. 그러나 NO 소거 활성으로 항염증 활성을 측정한 결과, 발효 후 농도 의존적으로 증가하는 것을 확인하였으며, L. plantarum로 발효한 발효액보다 L. brevis로 발효한 발효액의 항염증 활성이 더 높았음을 확인하였다. 작두콩 발효액의 항균 활성을 측정한 결과, 녹농균인 Pseudomonas aeruginosa, 포도상구균인 Staphylococcus sp., 대장균인 Escherichia coli에 대한 억제 효과가 우수하였으며, 세포 독성 측정 결과, 모든 실험군에서 대식세포인 RAW 264.7에 대한 독성을 보이지 않았다. 이와 같은 결과는 발효에 따른 항염증 및 항균 활성이 증가하였음을 보여주었으며, 이를 토대로 다양한 바이오 및 식품산업 분야에 효과적으로 활용될 수 있을 것으로 생각된다. 다만 추후에는 발효 조건의 확립과 항염증 및 항균 활성에 대한 정확한 메커니즘 연구가 필요할 것으로 판단된다.