








서 론
식혜는 우리나라의 대표적인 음청류의 하나로 기원과 유래는 알 수 없으나 엿기름을 일정시간 따뜻한 물로 추출하고, 쌀이나 찹쌀로 만든 고두밥과 혼합하여 당화시켜 제조하는 음료로, 각 가정의 명절 등의 행사에 떡과 함께 빼놓을 수 없는 귀중한 음료로 애용되어 왔다(Jeong 등, 2014). 감주 또는 단술로 불리기도 하는, 식혜는 곡물 전분이 엿기름의 amylase에 의한 당화작용으로 쌀 또는 찹쌀의 전분이 당화되면서 maltose, glucose 등의 감미와 식혜 고유의 풍미가 생성된다(Kim 등, 2007). 또한, 식혜는 비타민, 무기질 및 섬유질의 함량이 높은 식품 영양성이 우수한 한국 고유의 전통 음청류로 소화 개선 및 변비에 효과가 있다고 알려지면서 주춤했던 식혜의 시장이 최근 다시 성장하는 추세이다(Lee와 Yi, 2012). 특히 전통음료에 대한 관심이 증가하면서 기능성이 향상된 식혜에 관한 다양한 연구가 진행되고 있으며, 고혈압, 항암 및 항균활성이 우수한 녹차를 이용하여 제조한 식혜의 관능 및 저장성 향상(Park, 2006), coumarin, capillone, β-pinene 및 alkaloid 성분을 다량 함유하고 있는 인진쑥 추출물을 첨가하여 제조한 식혜의 항산화 활성의 향상(Song과 Hwang, 2016)과 같은 기능성이 우수한 향토 자원을 활용하여 제조한 식혜에 대한 연구가 활발히 진행되고 있다.
현미는 벼의 왕겨층만 벗겨낸 것으로 쌀겨층, 배, 배젖으로 구성되어 있으며, 백미에 비해 도정으로 인한 영양분의 손실이 적고 저장성이 높은 장점을 갖고 있다. 또한, 지방, 단백질, 비타민 및 식이섬유의 함량이 높고, γ-oryzanol, γ-aminobutyric acid(GABA)와 같은 다양한 기능성 물질이 많이 함유되어 있으며, 폴리페놀 화합물을 다량 함유하고 있어 항산화 활성, 혈압 및 지질 대사 개선, 면역력 증강 등의 기능성을 지녀 건강증진 효과 또한 우수하다(Kim 등, 2011). 하지만 백미의 섭취보다 영양학적으로는 우수하나 낮은 도정도로 인하여 소화율이 감소하고, 식미 또한 반감되는 단점도 함께 가지고 있다(Kim 등, 2006). 따라서 현미의 영양학적 가치와 기능성을 이용함과 동시에 소화력 향상을 위한 방법으로 현미의 가공을 통해 다양한 제품 개발에 이용하는 방법이 일반적으로 사용되고 있다. 특히 현미를 이용하여 막걸리를 제조한 결과, 유기산의 함량이 증가하고 유리아미노산 또한 증가하여 기능성이 증진되었다는 보고(Baek 등, 2013)가 있으며, 최근에는 미생물을 이용하여 발효 중 미생물이 분비하는 가수분해효소에 의한 전분, 단백질과 같은 고분자 물질의 분해로 소화력의 증진과 다양한 유기산, 올리고당, 단백질 등과 같은 각종 생리활성 물질을 공급하는 방법이 개발되고 있다(Kiers 등, 2000; Park와 Oh, 2005). 미생물에 의한 현미의 발효 방법으로는 일반적으로 곰팡이가 사용되나(Takasuga 등, 2004), 최근에는 유산균의 초기 생장을 도와주기 위해 다른 영양원을 첨가하거나 현미에 당화시킨 백미를 첨가하여 발효시킴으로써 장 기능에 도움을 주는 프로바이오틱스 유산균을 이용한 발효법이 보고되고 있다(Jeoun 등 1995; Kim 등, 2006).
일반적으로 김치, 치즈와 같은 발효 식품에 많이 존재하고 있는 유산균은 프로바이오틱스 균주로 변비 개선과 아토피와 같은 면역질환의 개선에 효과적인 것으로 알려져 있으며(Seo와 Lee, 2007), 항균 물질을 생성하여 유해균의 증식을 억제하고, 과민성 대장증후군 및 유당 불내증 개선에 대한 기능성이 우수하다(Seevedra 등, 2004). 이외에도 프로바이오틱스의 다양한 기능성이 보고되면서 고령화 및 식습관의 변화로 인하여 대사성 질환의 예방을 위한 소재로 프로바이오틱스가 각광을 받고 있으며, 사회적 트렌드를 반영하여 프로바이오틱스 유산균을 이용한 기능성 향상 제품들이 지속적으로 개발되고 있다. 약초 추출액과 유산균을 이용하여 젖산 발효를 통해 항산화 활성 및 유기산, 페놀 생성량 증가를 통한 기능성 증진 감주(Cho 등, 2008), 당근 추출물을 유산균 발효를 통한 침전물 억제 및 품질이 향상된 당근 주스(Jo 등, 2008), 전통발효 식품으로부터 분리한 유산균을 이용하여 제조한 요거트의 항산화 효과 향상(Park과 Lee, 2017) 등 다양한 연구 결과를 기반으로 기능성 제품이 출시되고 있다. 하지만 유산균이 식품에 사용되기 위해서는 무엇보다도 안전성, 기능성 및 기술적 요소에서의 검증이 필요하며, 최근에는 프로바이오틱스에 대한 규제 및 검증이 강화되고 있는 추세이다(Bang 등, 2012). 유산균은 무엇보다도 인간과 동물의 장내 환경에서 생존하기 위하여 위산, 담즙에 대한 내성을 지녀야 하며, 숙주의 장내에서 유익한 역할을 수행하기 위해서는 장내 세포 표면에 대한 부착능이 중요한 요소로 알려지고 있으며, 인돌, 아민과 같은 유해 대사산물이나 대장암의 원인인 β-glucoronidase와 같은 유해효소를 생성하지 않아야 한다(Cole 등, 1989; Granato 등, 1999; Henriksson 등, 1999).
따라서 본 연구에서는 현미 식혜 제조를 위하여 국내 전통발효 식품으로부터 유산균을 분리하고 프로바이오틱 특성 및 유해물질에 대한 생성 여부 조사를 통해 현미 식혜 제조용 균주를 선별하고, 이를 이용한 현미 식혜 제조 중 발효 특성 및 기능성을 검토하였다.
재료 및 방법
유산균 분리를 위한 시료는 전라북도 순창군에서 전통적인 방법으로 제조한 메주 시료 약 100여 종을 수집하여 사용하였으며, 각각의 시료 1 g을 9 mL의 멸균 생리식염수에 단계 희석 후 100 μL를 MRS(Difco™, MI, USA) 고체 배지에 도말하여 37°C에서 48시간 배양한 후 생성된 콜로니를 선택하여 순수 분리하였다. 유산균 분리주는 다음 연구를 위하여 10% skim milk(Difco™, MI, USA)에 재부유하고 −80°C에 보관하여 다음 연구에 사용하였다.
선별 유산균의 식품 내 식중독 유발 균주 중 하나인 Bacillus cereus를 대상으로 well diffusion 방법을 이용하여 측정하였다. B. cereus KCCM 40935 균주를 24시간 동안 Nutrient broth(NB, Difco™, MI, USA) 배지에 접종하여 30°C에서 배양한 후 배양액의 흡광도를 측정하여 일정한 농도(OD660 nm, 0.4)로 0.8% 한천을 함유한 NB(Difco™) soft 고체 배지에 첨가하여 평판 배지를 제조하였다. 활성 측정을 위한 유산균 시료는 MRS(Difco™) 액체 배지에 24시간 동안 37°C에서 진탕 배양한 후 13,000 rpm으로 10분간 원심분리한 후 상등액을 취하여 0.45 μm syringe filter(Sartorius, Frankfurt, Germany)로 제균하고 6 mm의 well에 각각의 상등액을 분주하였다. 항균 활성은 30°C에서 24시간 배양한 후 각각의 well 주변에 형성된 억제환의 크기를 측정하였다.
바이오제닉 아민의 분석은 1,000 mg/L의 바이오제닉 아민 전구체 tyrosine(Sigma-Aldrich, MO, US)과 histidine(Sigma-Aldrich, MO, US)이 함유된 MRS(Difco™) 액체 배지에 선별한 유산균 분리주를 각각 접종하고 37°C, 24시간 배양한 후 각각의 배양액을 13,000 rpm으로 30분간 원심분리하여 상등액을 회수하였다. 표준 시료는 0.01 N HCl(Sigma-Aldrich, MO, US)에 1 g/L가 되도록 제조한 후 0.1–100 mg/L의 농도로 단계 희석하여 검량선 작성을 위한 시료로 사용하였다. 앞서 준비한 각각의 시료와 표준 용액 0.5 mL에 0.25 mL의 내부표준용액 1,7-diaminoheptane(0.1 g/L, Sigma-Aldrich, MO, US)을 첨가하고, 0.25 mL의 포화 Na2CO3(Sigma-Aldrich, MO, US)와 0.4 mL의 1% dansyl chloride(Sigma-Aldrich, MO, US)를 첨가한 후 45°C에서 1시간 유도체화를 실시하였다. 유도체화 후 0.25 mL의 10% proline(Sigma-Aldrich, MO, US)을 첨가하여 남아있는 dansyl chloride(Sigma-Aldrich)를 제거하고, 2.5 mL의 ethyl ether(Samchun, Seoul, Korea)을 가하여 3분간 진탕한 후 상등액을 취하고 질소 농축기에서 완전히 증발시킨 후의 잔여물을 0.5 mL의 acetonitrile(Duksan, Seoul, Korea)에 녹인 후 0.45 μm syringe filter(Sartorius)로 여과하여 바이오제닉 아민 분석을 위한 시료로 사용하였다. 각각의 전처리 시료는 Seo 등(2018)의 방법에 따라 조사하였다.
선별 유산균의 용혈성 측정을 위한 5% blood 고체 배지는 5% sheep blood(MB cell, Seoul, Korea)를 MRS(Difco™)에 첨가하여 제조하였다. 각각의 선별 유산균을 접종하여 37°C에서 24시간 동안 배양한 후 균주 주위에 형성된 환의 형태를 통해 용혈성 여부를 판단하였다. 유해 대사산물 중 indole의 생성 여부는 각각의 선별 유산균을 0.8%의 tryptophan(Sigma-Aldrich, MO, US)을 함유한 MRS(Difco™) 액체 배지에 접종하고 37°C, 24시간 진탕배양한 후 indole reagent dropper(Difco™, MI, USA) 를 첨가하여 표면에 나타나는 색의 변화를 육안으로 관찰하여 indole의 생성 여부를 확인하였다. Phenyl-pyruvic acid 생성능은 phenylalanine agar(Fluka, Buchs, Switzerland) 고체 배지에 획선 도말한 후 37°C에서 24시간, 48시간 배양한 후 10%의 ferric-chloride(MB cell, Seoul, Korea) 3–4 방울을 떨어뜨리고 균주 주위의 색의 변화를 관찰하여 생성 여부를 확인하였다. 유해 효소의 하나인 β-glucuronidase는 tryptone bile X-glucuronide(TBX, Oxoid, Hampshire, UK) 배지에 획선 도말하여 37°C, 24시간 배양 후 색의 변화를 관찰하여 측정하였다.
시료의 항산화 활성 측정은 2,2-diphenyl-1-picryl-hydrazyl (DPPH, Sigma-Aldrich, MO, US)에 의한 free radical 소거능과 superoxide dismutase(SOD) 유사 활성능을 측정하여 확인하였다. 항산화 활성 측정을 위한 각각의 시료 배양액과 식혜를 13,000 rpm에서 10분간 원심분리 후 상등액을 취하여 측정하였다. Free radical 소거능은 100 μM DPPH ethanol 용액 180 μL에 각각의 시료 20 μL를 혼합하고, 암실 조건하에 30분간 반응시킨 후 UV/VIS spectrophotometer(SPECORD200, Analytik Jena, Jena, Germany)로 517 nm에서 측정한 후 아래 식에 대입하여 산출하였으며, 대조구로는 MRS(Difco™) 배지를 사용하였다.
SOD 유사 활성능은 SOD kit(Sigma-Aldrich, MO, US)를 이용하여 측정하였으며, 각각의 시료 20 μL에 WST working solution 200 μL와 enzyme working solution 20 μL를 혼합하고, 37°C에서 20분간 반응시킨 후 450 nm에서 흡광도를 측정하였다.
선별 유산균의 β-glucosidase 활성 측정은 1% esculin (Difco™, MI, USA)과 0.5% ferric ammonium citrate(Junsei, Tokyo, Japan)을 첨가한 MRS(Difco™) 배지에 접종하고 37°C에서 24시간 배양한 후 형성된 검정색 환의 크기를 측정하였다.
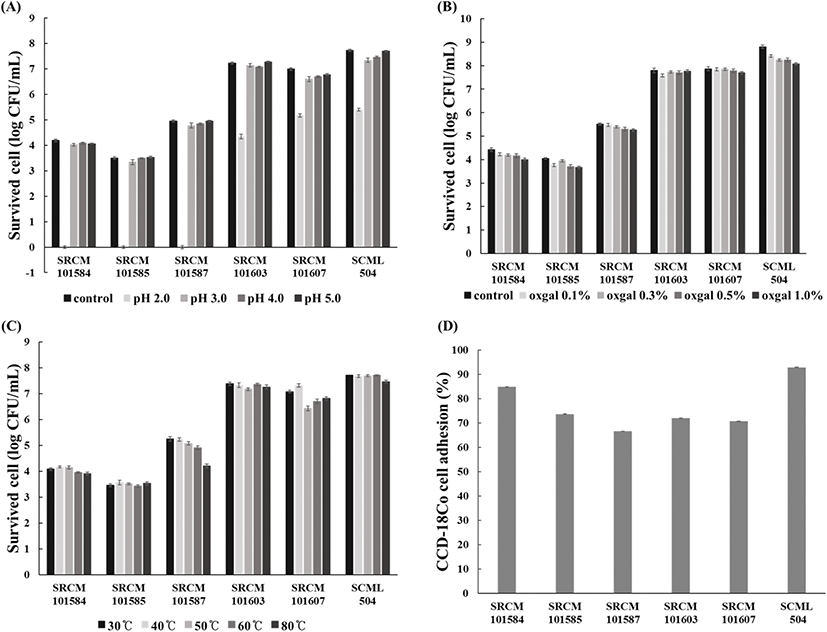
선별 균주의 프로바이오틱스 특성분석은 내산성, 내담즙성, 내열성 및 장내 부착능을 측정하여 확인하였다. 내산성은 각각의 선별 균주 배양액을 pH 2, 3, 4, 5로 조절한 MRS(Difco™) 액체 배지에 접종하고 37°C에서 3시간 배양한 후 각각의 생균수를 측정하였으며, 대조구로는 pH 조절을 하지 않은 MRS(Difco™) 배지에서 동일 조건하에 배양하여 생균수를 측정하여 사용하였다. 내담즙성은 각각의 선별 균주 배양액을 oxgall(Difco™, MI, USA)이 0.1%(w/v), 0.3%(w/v), 0.5% (w/v), 1.0%(w/v)로 함유된 MRS(Difco™) 액체 배지에 접종하고 37°C에서 3시간 배양한 후 균수를 측정하여 확인하였고, 대조구로 MRS(Difco™) 배지에서 동일 조건에서 배양한 생균수를 측정하여 사용하였다. 열에 대한 저항성은 MRS(Difco™) 액체 배지에 각각의 선별 유산균을 접종한 후 30°C, 40°C, 50°C, 60°C, 80°C에서 10분간 배양한 후 각각의 생균수를 측정하여 확인하였다. 또한, 장 상피 세포 부착능은 한국세포주은행으로부터 CCD-18Co 세포를 분양받아 Eom 등(2015)의 방법에 따라 측정하였다. CCD-18Co 세포를 24-well tissue culture plate(SPL Life Sciences Co. Ltd., Pocheon, Korea)에 2×105 cell/well이 되도록 37°C 혐기배양장치에서 24시간 배양하였다. 각각의 선별 균주를 multiplicity of infection(MOI) 10의 농도로 처리하여 45분간 감염시킨 후 PBS buffer(Sigma-Aldrich, MO, US)로 3회 세척하였으며, 200 μL의 0.1% triton X-100(Sigma-Aldrich, MO, US)를 처리하여 CCD-18Co 세포에 부착된 균을 회수하고 MRS(Difco™) 고체 배지에 도말하여 생균수를 측정하여 아래 식에 대입하여 산출하였다.
최종 균주의 효소 활성은 API ZYM kit(Biomerieux, Marcy-I’Etoile, France)를 이용하여 측정하였다. MRS (Difco™) 액체 배지에 선별 균주를 접종하고 37°C에서 24시간 배양한 후 균체를 회수하여 멸균수로 3회 세척하고 멸균수에 현탁하여 시료를 준비하였다. 5 mL suspension medium(Biomerieux, Marcy-I’Etoile, France)에 탁도 5–6 Mcfarland(Biomerieux, Marcy-I’Etoile, France)로 조정하고, 현탁액을 ZYM kit의 각 큐플에 접종하여 37°C에서 4시간 배양한 후 표현 활성 증가와 용해를 도와주는 ZYM A, B 시약을 각각 큐플에 떨어뜨려 5분간 반응시킨 후 색의 변화를 관찰하여 효소 활성 정도를 측정하였다.
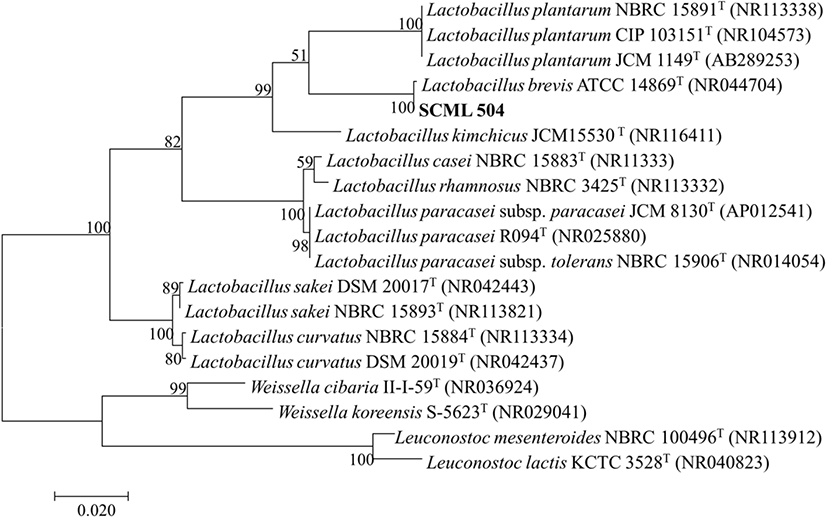
최종 선별 균주의 동정을 위하여 16S rRNA 유전자 분석을 진행하였다. 최종 선별 균주의 균체를 회수하여 ZR Fungal/Bacterial DNA Miniprep kit(Zymo Research Corp., CA, USA)로 DNA를 추출한 후 Ryu 등(2018)의 방법에 따라 PCR로 유전자 단편을 증폭하고 정제하여 염기서열을 ㈜마크로젠에 의뢰하여 분석하였다. 분석 결과는 National Center for Biotechnology Information(NCBI, Bethesda, MD, USA)의 BLAST를 이용하여 서열 일치도가 높은 표준 균주의 16S rRNA 염기서열을 확보하여 계통도를 작성하였다. 계통도 분석은 Neighbor-joining 알고리즘(Kumar 등, 2004)을 이용하여 분석하였으며, 계통수의 각 분지에 대한 신뢰도 산출을 위하여 bootstrap 분석을 1,000회 반복 수행하여 견고성을 확인하였다.
식혜 제조를 위하여 정읍산 참좋은 일반현미, 함양 농협의 하늘가애 엿기름 제품을 구입하여 사용하였다. 식혜 제조를 위한 엿기름 추출물은 물 2 L에 엿기름 200 g을 첨가하고 40°C에서 130 rpm으로 3시간 진탕 추출하였으며, 고두밥은 현미 20 g을 1시간 수침하고 121°C에서 10분간 고압증기 멸균하여 제조하였다. 제조한 고두밥 20 g에 엿기름 추출액 200 mL를 첨가하고 60°C에서 6시간 당화한 후 100°C에서 1시간 가열하고, 30°C 이하로 냉각하여 제조하였다. 식혜 유산균 발효액은 최종 선별 균주를 MRS (Difco™) 액체 배지에서 37°C, 24시간 배양한 후 식혜의 1%에 해당하는 균체를 회수하여 멸균수로 3회 세척 후 식혜에 접종하고 37°C에서 정치 발효하였다.
유산균 발효 식혜의 발효 특성 분석은 발효 0시간에서 72시간까지 24시간 단위로 시료를 회수하여 유산균의 생균수, pH 및 당도의 변화를 측정하여 조사하였다. 유산균의 균체 성장은 발효 식혜 1 mL를 취하여 멸균 생리식염수 9 mL에 단계희석하고 MRS(Difco™) 고체 배지에 각각 시료 100 μL를 도말하여 생균수를 측정하였다. 발효 시간에 따른 pH 및 당도의 변화는 각각 회수한 시료를 pH meter(S220, Mettler Toledo Gmbh, Schwarzenbach, Switzerland)와 디지털 당도계(ATAGo Co. Ltd., Tokyo, Japan)를 이용하여 측정하였다.
발효 시간에 따른 유산균 발효 식혜의 유리당 및 유기산의 분석은 각각 24시간 단위로 회수한 시료를 13,000 rpm에서 10분간 원심분리하고, 0.45 μm syringe filter(Sartorius)로 여과 후 HPLC(Agilent 1200 series, Agilent Technologies, Santa Clara, CA, USA)를 이용하여 측정하였다. 분석을 위한 column은 Aminex HPX-87H(300×7.8 mm, Bio-RAD, Hercules, CA, USA)를 사용하여 측정하였으며, 이동상은 5 mM sulfuric acid(Sigma-Aldrich, MO, US), 유속은 0.6 mL/min, 검출기는 DAD detector 210 nm(Agilent Technologies, Santa Clara, CA, USA)와 RID detector(Agilent Technologies, Santa Clara, CA, USA)를 이용하여 동시 분석하였다.
유산균 발효 식혜의 발효 시간에 따른 유리아미노산의 측정은 아미노산 분석기(L-8900, Hitachi High-Technologies, Tokyo, Japan)를 이용하여 측정하였다. 24시간 단위로 회수한 발효물을 3,000 rpm에서 10분간 원심분리한 후 각각 상등액 2 mL에 5% trichloro-acetic acid(TCA, Sigma-Aldrich, MO, US) 2 mL를 첨가하고 4°C에서 1시간 반응시킨 후 13,000 rpm에서 10분간 원심분리를 실시하였다. 상등액 500 μL에 500 μL의 0.02 N HCl(Sigma-aldrich)을 첨가하고, 0.45 μm syringe filter(Sartorius)로 여과한 후 분석에 사용하였으며, 기기 분석 조건은 Jeong 등(2014)의 방법에 따라 분석하였다.
결과 및 고찰
식혜 발효에 적합한 유산균의 분리를 위해 전라북도 순창군의 일반 가정집에서 다양한 방식으로 제조된 메주 100여 종을 수집하여 균원 시료로 사용하였다. 수집한 각각의 시료는 단계 희석하여 MRS 고체 배지에 배양한 후 집락의 형태, 색 등이 상이한 분리주 260종을 분리하였으며, 식혜 발효에 사용하기 위한 유산균 후보 소재로 활용하였다.
B. cereus는 토양, 공기 중에 부유하는 미생물로 식품에서 쉽게 오염되어 부패를 유발시키고 체내 과량 섭취할 경우에는 구토와 설사 등의 증상을 발병시키게 된다. 최근에는 유산균이 생산하는 유기산과 박테리오신 등과 같은 항균 물질이 식품에 발생하는 유해균을 효과적으로 억제하는 것이 알려지면서 유산균의 식품 유해미생물에 대한 항균 효과가 중요한 선별 요소 중 하나로 제시되고 있다(Enan 등, 1996). 따라서 앞서 확보한 분리주를 대상으로 항균 활성을 측정한 결과 B. cereus KCCM 40935에 대하여 10 mm 이상의 활성을 갖는 균주 6종을 1차 선별하였으며, 특히 SCML 504는 15 mm의 활성을 지녀 다른 균주에 비해 식중독 유발균 B. cereus에 대한 효과가 가장 우수함을 확인하였다(Table 1).
일반적으로 바이오제닉 아민은 아미노산 탈탄산 효소를 생성하는 미생물에 의해 발효 과정에서 생성되는 저분자 질소 화합물로 고농도로 섭취할 경우 두통, 두드러기, 호흡곤란 등이 발생하게 되어 발효 식품, 특히 한국의 전통 발효 식품에서 그 유해성이 드러나기 시작하면서 사회적으로 관심이 증가하고 있는 물질의 하나이다(Seo 등, 2018). 따라서, 발효에 사용되는 유산균의 바이오제닉 아민 생성 여부는 식품 소재로 사용하기 위한 대표적인 안전성 관련 지표 중 하나이며, Lim(2017)에 따르면 유산균이 생산하는 물질에 의해 바이오제닉 아민을 생산하는 미생물이 억제되거나 탈탄산 효소의 활성을 억제한다고 보고하고 있어, 식혜 발효에 사용하기 위한 유산균 후보 소재의 바이오제닉 아민 생성 여부를 측정한 결과 SRCM 101607을 제외하고, 모든 선별 균주에서 생성하지 않거나 인체에 무해한 수준으로 매우 적은 양이 생성되는 것을 확인하였다(Table 1).
유산균을 식품에 적용하기 위해서는 항균력, 바이오제닉 아민 생성 여부 외에도 병원성 및 알레르기에 대하여 인체에 유해한 물질이 생성되지 않아야 하기 때문에, 용혈성, 유해 대사산물 및 유해효소에 대한 생성 여부를 측정하였다. 용혈성은 크게 적혈구의 불완전 용해로 인하여 세균 집락이 녹색의 불투명한 환을 생성하는 α-hemolysis, 적혈구를 완전 용해시켜 황색의 환을 생성하는 β-hemolysis와 용혈 현상이 발생하지 않는 γ-hemolysis로 구분되며, 6종의 선별 균주 모두 용혈성이 발생하지 않아, Park 등(2017)의 결과와 동일하게 선별 균주 모두 용혈성에 대한 안전성이 우수함을 확인하였다(Table 1). 또한, 암 유발 물질 중 하나인 indole과 phenyl-pyruvic acid 역시 선별 균주 모두 생성하지 않음을 확인하였으나, 간에서 benzopyrene 등 β-glucoside 배당체 화합물과 glucuronic acid 화합물이 glucuronic acid conjugate로 무독화되어 장으로 보내졌을 때 이 결합을 끊어 주어 발암 원인을 제공하는 것으로 보고(Nanno 등, 1986)된 β-glucuronidase의 경우 SRCM 101607 균주에서만 검출되었고, 나머지 5종의 균주는 독성 물질을 생성하지 않는 것을 최종 확인하였다(Table 1).
일부 유산균은 활성 산소 소거를 통해 체내 산화적 손상을 유발하여 유전자의 손상과 발암을 유발하는 항산화 효과를 가지며, 이로 인해 노화를 억제하는 중요한 효과를 나타낸다고 보고되고 있다(Seo 등, 2018). 따라서 앞서 선별한 6종의 유산균을 대상으로 DPPH에 대한 항산화 활성을 측정한 결과 선별 유산균 모두 20% 이상의 항산화 활성을 갖는 것을 확인하였다(Table 1). 이러한 결과는 Seo 등(2018)의 보고에서 유산균 8종이 10–20% 항산화 활성을 갖는다고 보고한 것과 유사한 결과를 보이고 있다. 따라서 선별 균주 6종 모두 활성산소의 억제를 통한 식품, 사료 등 다양한 분야에 항산화제로 사용될 수 있는 가능성을 확인하였다. 또한, Cho 등(2008)은 약초 추출액과 유산균을 이용하여 젖산 발효 시 항산화 활성이 증가한 감주를 제조하였다고 보고하고 있어, 현미 또한 유산균 발효를 통해 더욱더 항산화 활성이 증가할 것으로 판단하였다.
β-Glucosidase는 유산균이 생성하는 효소 중 하나로 배당체 화합물을 유리시키는 특이성을 갖고 있어 식품 산업에 다양한 분야에 활용되고 있으며, 그 중 대두의 isoflavone을 위산과 장내 미생물의 β-glucosidase에 의하여 비배당체로 유리시킴으로써 장내 흡수율을 향상시키는 역할을 한다고 보고되고 있다(Yang 등, 2007). 선별 유산균의 β-glucosidase 활성을 측정하기 위하여 esculine을 첨가한 MRS-EI 고체 배지로 확인한 결과, SRCM 101603, SRCM 101607, SCML 504 균주가 높은 활성을 갖고 있었다(Table 1). 따라서 이들 3종을 이용하여 식품 제조에 사용할 경우 체내 흡수가 용이한 형태로 배당체 화합물을 유리시킬 수 있는 기능성을 갖는 것을 확인하여 식품 제조 시 응용할 수 있는 분야가 확대될 가능성을 확인하였다.
유산균을 프로바이오틱스로 이용하기 위해서는 위산과 담즙이 존재하는 환경에서 생존할 수 있는 능력을 지녀야 한다. 따라서 산에 대한 생존능을 확인한 결과, pH 3.0, 4.0, 5.0에서는 모든 균주가 대조구인 초기 투입 균수와 유사한 생존율로 존재하는 것을 확인하였다. 특히 위산의 pH와 유사한 2.0에서도 SRCM 101603은 60.08%, SRCM 101607은 73.94%, SCML 504는 69.80%로 산에 대한 저항성이 매우 우수함을 확인하였으며(Fig. 1A), 담즙에 대한 저항성 또한 선별 균주 6종 모두 장내 담즙 농도(0.3%)보다 높은 0.5% 이상의 oxgall에서도 약 90%의 높은 생존율을 갖는 것을 확인하였다(Fig. 1B). 유산균은 일반적으로 내산성이 약하다고 보고되어 있지만, L. brevis는 일부 높은 내산성을 지닌다는 보고(Park, 1995)와 일치하는 결과를 얻었으며, 내산성이 우수한 유산균이 담즙에서도 우수한 내성을 지닌다는 보고(Shin 등, 1999)와도 일치하는 결과를 확인하였다. 또한, 일반적으로 식품가공 공정의 경우 살균 또는 제품 제조를 위하여 60–80°C의 열처리 공정이 이루어지기에 열에 대한 안전성도 유산균의 경우 중요한 요소 중 하나이다. 따라서 선별 균주의 내열성을 측정한 결과에서도 80°C에서 6종의 선별 균주 모두 95% 이상의 높은 생존율을 갖는 것을 확인하였다(Fig. 1D). CCD-18Co 세포를 이용한 장내 상피세포 부착능을 확인한 결과 또한 6종의 선별 균주 모두 66% 이상의 부착능을 보였으며, 특히 SRCM 101584, SCML 504 균주가 84.93%와 92.93%로 가장 우수한 부착능을 보였다(Fig. 1C). Park 등(2017)은 김치에서 분리한 L. plantarum의 프로바이오틱 특성 분석 결과, 내담즙은 70%의 생존율을 보였고, 대조구로 사용한 L. rhamnosus GG의 경우 약 78%의 생존율을 보였고, HT-29 세포를 이용한 부착능 실험에서는 약 20%의 부착율을 보여 프로바이오틱스 활성이 우수하다고 보고하였다. 이러한 결과를 고려하면 앞서 선별한 6종의 유산균은 내산, 내담즙, 내열성에 대한 안정성이 우수하며, CCD-18Co 세포에 대한 부착능도 우수하여 프로바이오틱스 소재로의 활용 가능성이 매우 우수한 균주로 확인하였다. 최종적으로 프로바이오틱스 특성 및 기능성 분석 결과를 종합하여 가장 우수하며, 활용 범위가 넓은 SCML 504 균주를 최종 균주로 선정하고, 유산균 식혜 제조를 위한 유산균 소재로 결정하였다.
최종 선별 균주인 SCML 504 균주의 효소 생산능을 확인하기 위하여 API ZYM kit를 이용하여 분석한 결과, 지질에 대한 가수분해 효소인 esterase 및 esterase lipase에 대한 활성을 갖고 있었으며, protease 계열의 효소에도 활성을 갖는 것으로 확인되었다. 또한, raffinose나 stachyose 등을 분해하는 것으로 알려진 α-galactosidase에 대한 활성을 갖고 있었으며, β-galactosidase에 대해서도 활성을 나타내어 유당 불내증에 대한 단점도 보완할 수 있는 균주로 확인되었다(Table 2). 전반적인 효소 활성 측정 결과, SCML 504 균주는 일반적으로 보고된 L. brevis 균주와 유사한 효소 생산을 갖는 것으로 확인되었으나, Yang 등(2018)이 보고한 L. brevis SBB07 균주와는 다르게 α-galactosidase와 esterase에 대한 활성을 갖고 있는 것을 기반으로 일부 효소 생산 능력에 대하여는 차이가 있는 것을 확인하였다.
16S rRNA 유전자 염기서열 분석을 통해 최종 선별한 SCML 504 균주의 동정 결과 L. brevis로 확인되었으며, eztaxon serve에서 표준 균주와 상동성을 비교분석한 결과를 바탕으로 phylogenetic tree를 작성하였다(Fig. 2). 동정 결과를 기반으로 최종 선별한 균주를 Lactobacillus brevis SCML 504로 명명하였으며, 한국미생물보존센터(Korean Culture Center of Microorganisms, KCCM)에 Lactobacillus brevis KCCM 12526P로 기탁하였다.
현미를 이용하여 식혜를 제조한 후 최종 선별한 유산균 SCML 504 균주로 발효하여 유산균 발효 식혜를 제조하였다. 제조한 유산균 발효 현미 식혜의 발효 특성을 분석하기 위하여 먼저 균체 성장을 조사한 결과, 접종 후 24시간까지 급격히 균수가 증가하기 시작하여 72시간까지 서서히 증가하였다(Table 3). pH 또한 균체 성장과 동일한 경향으로 발효 24시간에 6.12에서 3.92로 급격하게 감소하고, 이후에는 서서히 감소하는 경향을 보였다. 당도의 경우에는 초기 8.0 °Brix에서 발효 시간에 따라 서서히 감소하기는 하였으나, 균체 성장과 pH의 변화와 같이 발효 24시간에 급격히 감소하거나 증가하는 경향을 나타내지 않음을 확인하였다. 물론 아직까지 유산균을 이용한 발효 식혜에 대한 연구는 전무한 실정이나 유산균 발효 식혜의 경우, pH가 균체 성장과 반대의 경향을 나타내게 되고, 젖산과 같은 물질의 생성으로 인해 pH가 감소하게 되어 외부 오염균의 성장이 어려운 환경을 형성하게 된다. 이로 인하여 유산균 발효 식품의 경우, 저장성을 향상시키는 기능을 수행한다고 보고되고 있어 현미 유산균 발효 식혜의 경우 발효에 의한 저장성도 일부 향상될 수 있을 것으로 판단하였다(Kumar 등, 2004; Yang 등, 2007).
천연물 또는 농산물을 항산화 효과를 갖는 유산균으로 발효한 결과, 최종 산물에서 항산화 효과가 일부 향상된다고 보고(Cho 등, 2008; Park과 Lee, 2017)에 따라, 현미 식혜의 유산균 발효를 통해 항산화 활성의 변화를 확인하였다. DPPH 라디컬 소거능 측정 결과 유산균 발효를 진행하지 않은 식혜는 66.42%의 활성을 나타내었으나, 발효 후 72.27%로 소폭 증가한 것을 확인하였다. 또한, SOD 활성을 측정한 결과에서도 발효 전 현미 식혜의 69.25%에서 발효 후 74.98%의 활성을 갖는 것으로 확인되어, SOD 및 DPPH 라디컬 소거능이 유사한 수치로 소폭 향상되는 것을 확인할 수 있었다(Table 3). 한방 감주를 젖산 발효한 후 DPPH 라디컬 소거능을 측정한 결과에서 한방 감주가 발효 시간에 따라 76.6–75.7%로 감소하기는 하였으나 발효 전 0시간과 비교하여 큰 변화를 보이지는 않았으며(시판 감주 DPPH 라디컬 소거능, 14.4%), 이는 젖산 발효 중 생성된 유기산에 의한 영향으로 보고하고 있어(Cho 등, 2008), 현미 유산균 발효 식혜의 경우에도 경향이 소폭 증가하여 다르기는 하나 약 70%의 수준으로 큰 변화를 보이지는 않고 일정 수준을 유지하고 있어 동일한 영향으로 항산화 활성이 크게 증가하거나 감소하지 않은 것으로 판단하였다.
식혜는 쌀과 같은 곡물을 엿기름의 amylase에 의해 당화를 거쳐 단맛을 내는 한국의 대표적인 발효 음료이다. 따라서, 식혜를 제조 후 선별 유산균을 이용하여 발효 시간에 따른 유리당 함량의 변화를 측정한 결과, 발효 초기 glucose와 fructose의 함량이 8,403.07 mg/L와 2,068.12 mg/L에서 발효 48시간 이후 5,369.06 mg/L와 1,180.23 mg/L로 급격히 감소하였고 이후 72시간까지는 큰 변화가 없이 일정 수준을 유지하였다. 하지만, sucrose의 함량은 발효 초기 5.99 mg/L에서 72시간 후 4.60 mg/L로 큰 변화를 나타내지 않았다(Table 3). 이는 유산균의 경우 발효 시간에 따라 일반적으로 glucose, fructose와 같은 6탄당을 주로 이용하여 lactose 등의 이당류 또는 ribose 등의 오탄당을 대사에 이용하여 균체 성장 및 이차 대사산물을 생성한다는 보고(Shin 등, 2017)와도 일치하는 결과를 확인하였다.
현미 식혜의 유산균 발효 시간에 따른 유리아미노산의 함량을 측정한 결과, glutamic acid의 경우 발효 전 24.92 mg/L에서 발효 24시간 후 5.51 mg/L로 급격히 감소한 후 72시간까지는 유지되었으며, glutamic acid를 전구물질로 사용한 γ-aminobutyric acid(GABA)의 함량은 발효 전 24.57 mg/L에서 발효 72시간에 43.15 mg/L로 약 2배가 증가하였다. 일반적으로 중추신경계 신경전달물질의 하나로 알려진 GABA는 비 단백질성 아미노산으로 심혈관 기능의 조절, 뇌 신호 체계의 균형 유지 및 대장암 세포 증식 억제 효과 등 다양한 기능성에 대한 연구가 진행되어 그 효과가 알려져 있으며(Ryu 등, 2017), GABA의 생산량 증가를 위한 방법으로 식품 등 제조 시 유산균 발효를 통하여 GABA 함량의 향상을 통해 기능성을 증진 시키는 연구가 진행되고 있다(Hwang 등, 2018). 또한, 식품의 맛을 좌우하는 유리아미노산의 경우, 단맛을 나타내는 alanine은 발효시간이 경과함에 따라 큰 변화는 없었으나 11 mg/L 가량 감소하였고, glycine의 경우 7 mg/L가 증가하였다. 쓴맛을 내는 leucine의 경우에도 큰 변화는 없이 7 mg/L 감소한 수치로 발효에 따라 단맛과 쓴맛에 관여하는 유리아미노산의 함량 변화에는 큰 영향을 주지 않음을 확인하였다(Table 4).
최근 고령화 시대에 들어서면서 사람들이 식품 및 보조식품의 선택에 기존 맛 뿐만 아니라 기능성에 대한 부분도 고려의 대상이 되고 있다. 특히 유산균에 대한 사회적 관심이 급증하면서 이를 이용한 식품 응용에 대한 연구가 활발히 이루어지고 있는 실정이다. 따라서 본 연구에서는 전통 발효 식품으로부터 항균활성, 항산화 및 프로바이오틱스 활성을 가지며, 유해 물질을 생성하지 않는 Lactobacillus brevis SCML 504 균주를 선별하였으며, 이를 이용해 지역 농특산 작물 중 하나인 현미를 이용한 유산균 발효 기능성 식혜를 제조하기 위한 연구를 진행하였다. 최종적으로 SCML 504를 이용하여 제조한 현미 유산균 발효 식혜의 경우 기본적인 맛과 풍미에 큰 변화는 없지만, 기능성 물질로 알려진 GABA의 함량이 2배가량 증가하여 향후 GABA의 생산성 증가 등을 통한 기능성 향상을 목적으로 식혜를 제조할 경우, 기능성 음료로서도 많은 가능성을 갖는 음료가 될 것이라 판단된다.